

Force transduction by Triton cytoskeletons
- PMID: 11839769
- PMCID: PMC2174068
- DOI: 10.1083/jcb.200110068
Force transduction by Triton cytoskeletons
Abstract
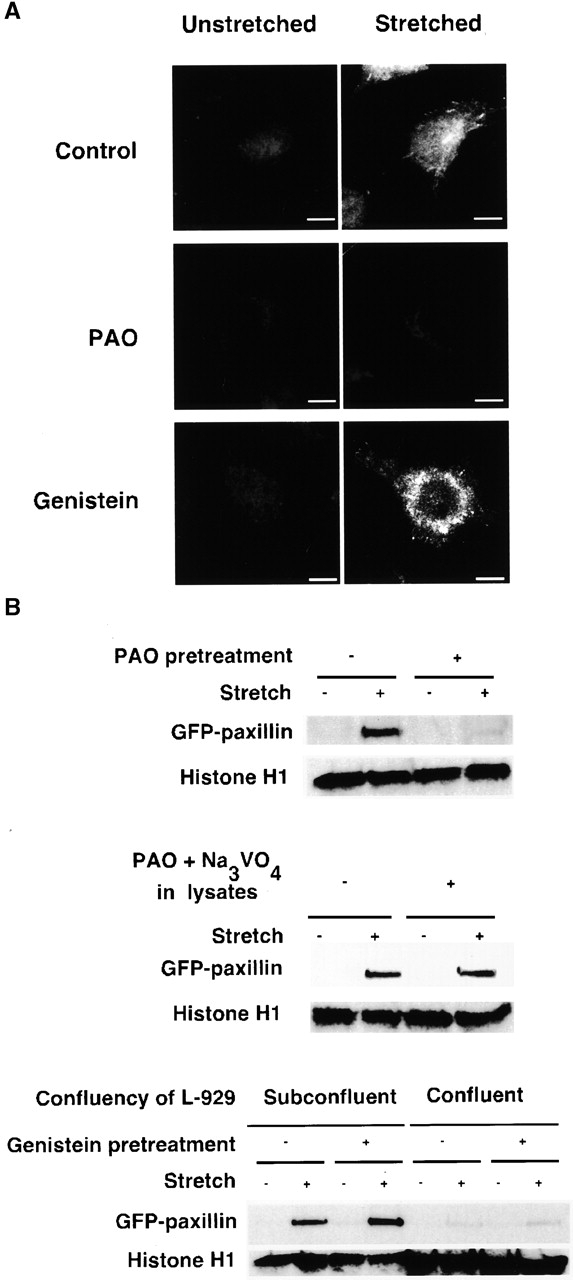
Force-initiated signal transduction can occur either via membrane-based ionic mechanisms or through changes in cytoskeletal-matrix linkages. We report here the stretch-dependent binding of cytoplasmic proteins to Triton X-100 cytoskeletons of L-929 cells grown on collagen-coated silicone. Triton X-100-insoluble cytoskeletons were stretched by 10% and incubated with biotinylated cytoplasmic proteins. Analysis with two-dimensional gel electrophoresis showed stretch-dependent binding of more than 10 cytoplasmic protein spots. Bound cytoplasmic proteins were purified by a photocleavable biotin tag and stretch-dependent binding of paxillin, focal adhesion kinase, and p130Cas was found, whereas the binding of vinculin was unchanged and actin binding decreased with stretch. Paxillin binding upon stretch was morphologically and biochemically similar in vitro and in vivo, that is, enhanced in the periphery and inhibited by the tyrosine phosphatase inhibitor, phenylarsine oxide. Thus, we suggest that transduction of matrix forces occurs through force-dependent conformation changes in the integrated cytoskeleton.
Figures
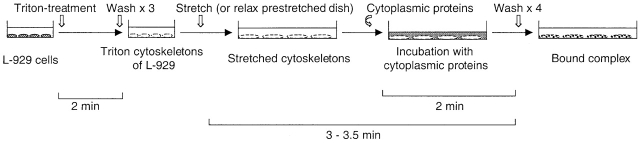
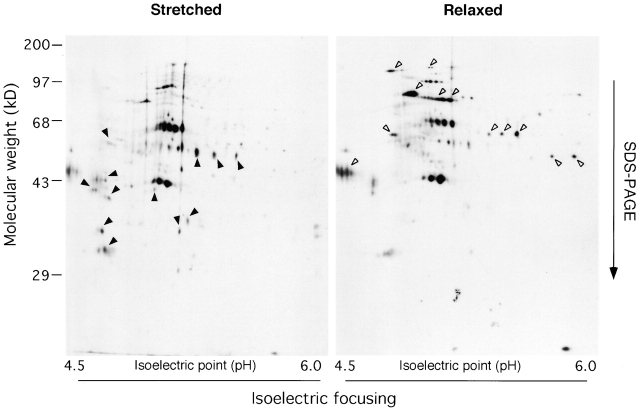
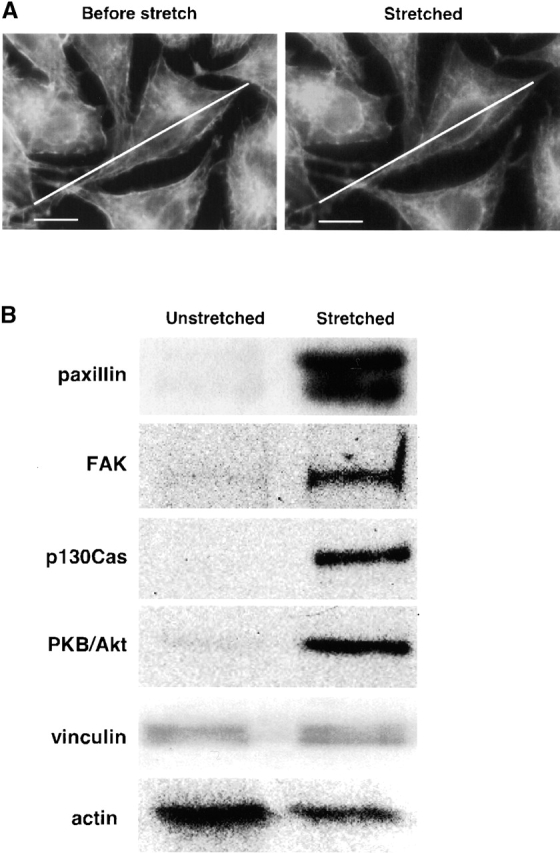
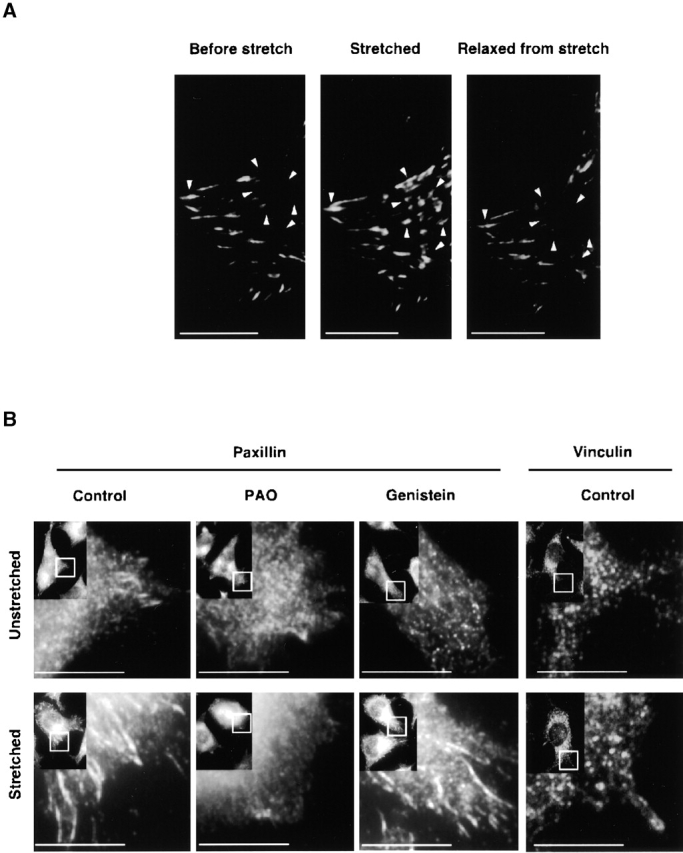
References
-
- Akiyama, T., J. Ishida, S. Nakagawa, H. Ogawara, S. Watanabe, N. Itoh, M. Shibuya, and Y. Fukami. 1987. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J. Biol. Chem. 262:5592–5595. - PubMed
-
- Balaban, N.Q., U.S. Schwarz, D. Riveline, P. Goichberg, G. Tzur, I. Sabanay, D. Mahalu, S. Safran, A. Bershadsky, L. Addadi, and B. Geiger. 2001. Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat. Cell Biol. 3:466–472. - PubMed
-
- Bang, O.S., E.J. Kim, J.G. Chung, S.R. Lee, T.K. Park, and S.S. Kang. 2000. Association of focal adhesion kinase with fibronectin and paxillin is required for precartilage condensation of chick mesenchymal cells. Biochem. Biophys. Res. Commun. 278:522–529. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
