

Integration sites for genetic elements in prokaryotic tRNA and tmRNA genes: sublocation preference of integrase subfamilies
- PMID: 11842097
- PMCID: PMC100330
- DOI: 10.1093/nar/30.4.866
Integration sites for genetic elements in prokaryotic tRNA and tmRNA genes: sublocation preference of integrase subfamilies
Abstract
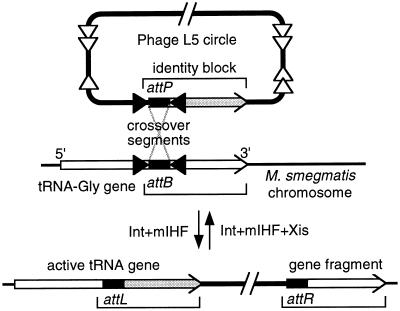
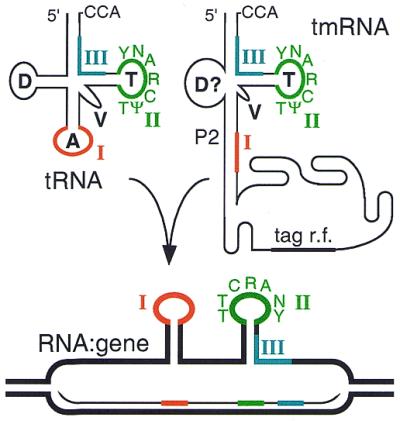
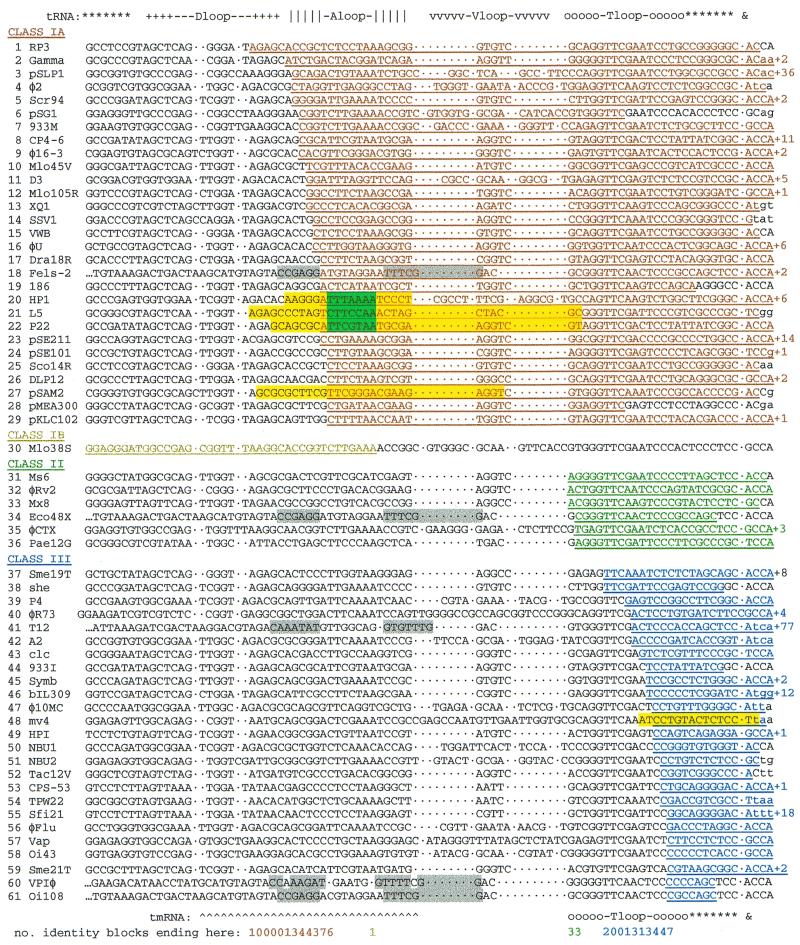
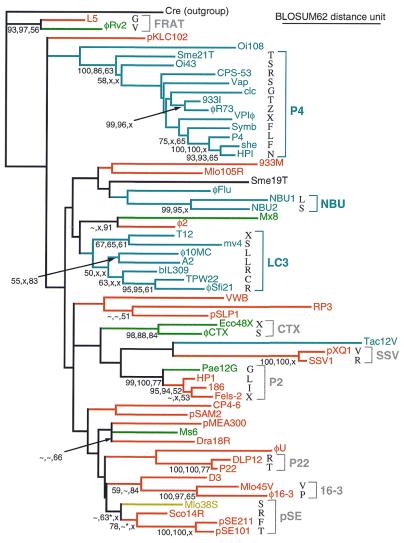
Most classical integrases of prokaryotic genetic elements specify integration into tRNA or tmRNA genes. Sequences shared between element and host integration sites suggest that crossover can occur at any of three sublocations within a tRNA gene, two with flanking symmetry (anticodon-loop and T-loop tDNA) and the third at the asymmetric 3' end of the gene. Integrase phylogeny matches this classification: integrase subfamilies use exclusively either the symmetric sublocations or the asymmetric sublocation, although tRNA genes of several different aminoacylation identities may be used within any subfamily. These two familial sublocation preferences imply two modes by which new integration site usage evolves. The tmRNA gene has been adopted as an integration site in both modes, and its distinctive structure imposes some constraints on proposed evolutionary mechanisms.
Figures
References
-
- Azaro M.A. and Landy,A. (2002) λ Integrase and the λ Int family. In Craig,N.L., Craigie,R., Gellert,M. and Lambowitz,A. (eds), Mobile DNA II. ASM Press, Washington DC, 118–148.
-
- Campbell A.M. (1962) Episomes. Adv. Genet., 11, 101–145.
-
- Weisberg R.A. and Landy,A. (1983) Site-specific recombination in phage lambda. In Hendrix,R.W., Roberts,J.W., Stahl,F.W. and Weisberg,R.A. (eds), Lambda II. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 211–250.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
