

Correlating protein footprinting with mutational analysis in the bacterial transcription factor sigma54 (sigmaN)
- PMID: 11842114
- PMCID: PMC100328
- DOI: 10.1093/nar/30.4.1016
Correlating protein footprinting with mutational analysis in the bacterial transcription factor sigma54 (sigmaN)
Abstract
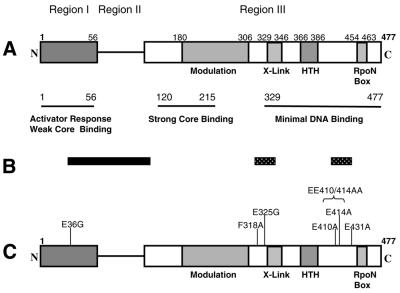
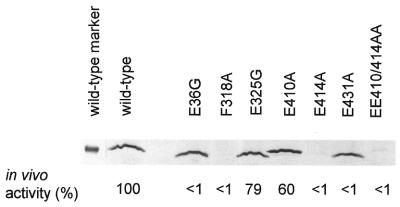
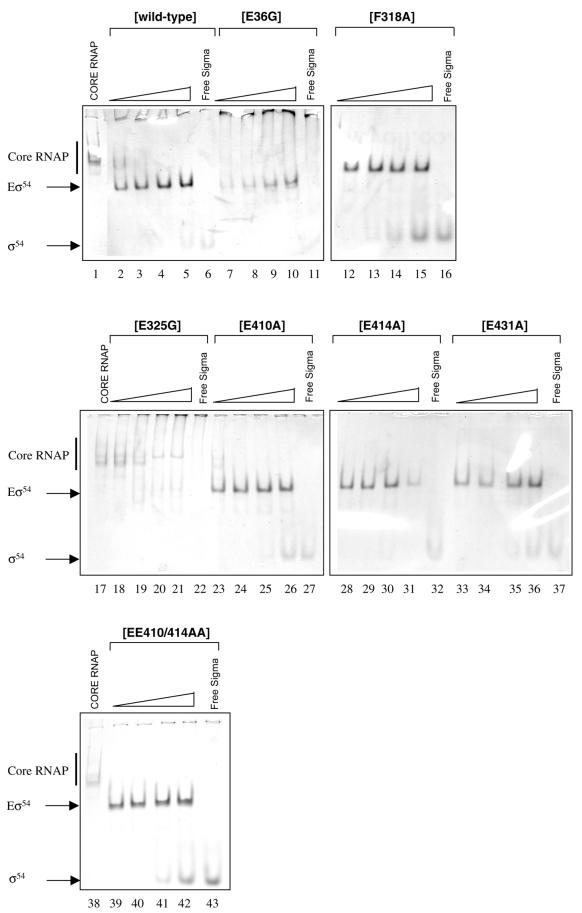
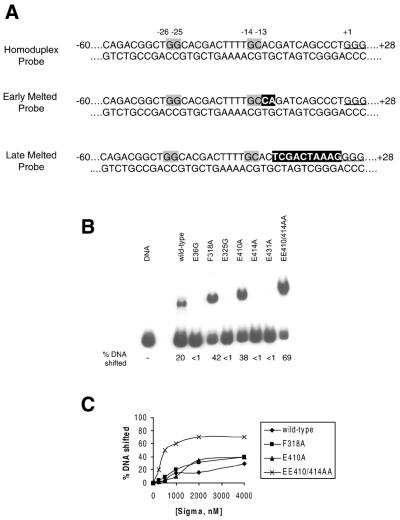
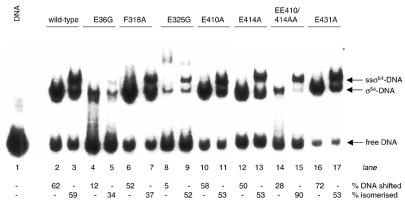
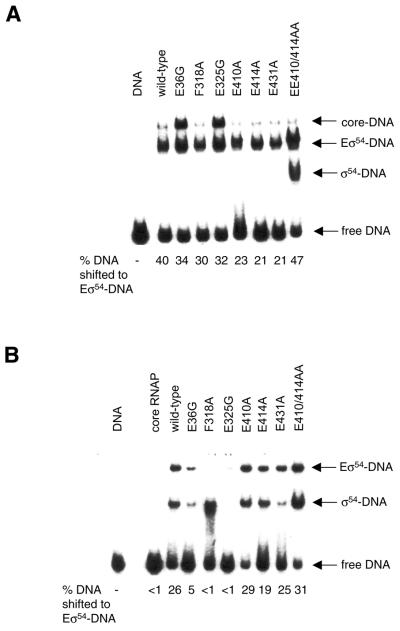
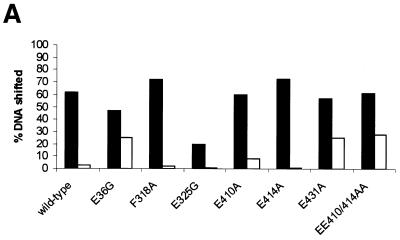
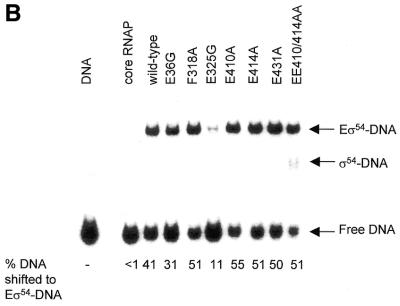
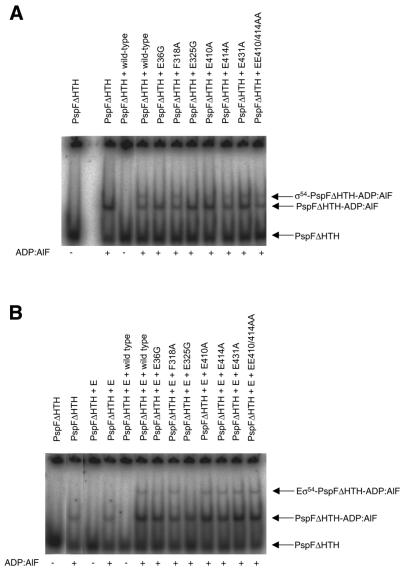
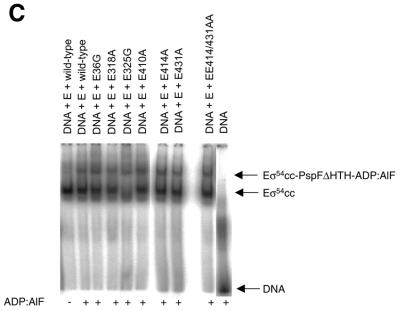
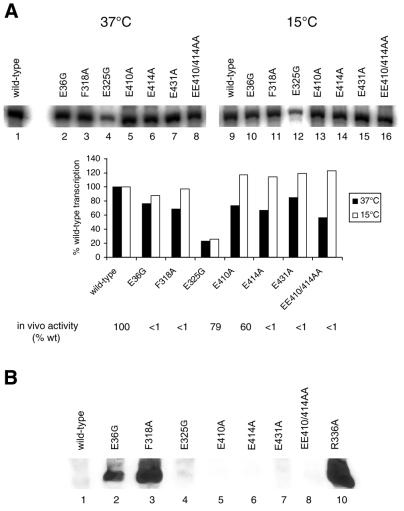
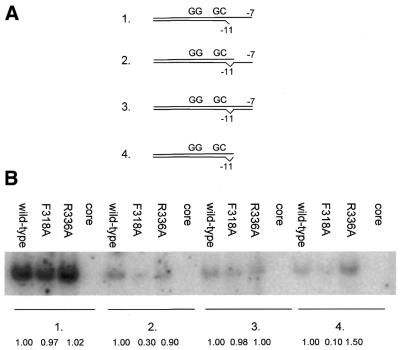
Protein footprints of the enhancer-dependent sigma54 protein, upon binding the Escherichia coli RNA polymerase core enzyme or upon forming closed promoter complexes, identified surface-exposed residues in sigma54 of potential functional importance at the interface between sigma54 and core RNA polymerases (RNAP) or DNA. We have now characterised alanine and glycine substitution mutants at several of these positions. Properties of the mutant sigma54s correlate protein footprints to activity. Some mutants show elevated DNA binding suggesting that promoter binding by holoenzyme may be limited to enable normal functioning. One such mutant (F318A) within the DNA binding domain of sigma54 shows a changed interaction with the promoter regulatory region implicated in transcription silencing and fails to silence transcription in vitro. It appears specifically defective in preferentially binding to a repressive DNA structure believed to restrict RNA polymerase isomerisation and is largely intact for activator responsiveness. Two mutants, one in the regulatory region I and the other within core interacting sequences of sigma54, failed to stably bind the activator in the presence of ADP-aluminium fluoride, an analogue of ATP in the transition state for hydrolysis. Overall, the data presented describe a collection sigma54 mutants that have escaped previous analysis and display an array of properties which allows the role of surface-exposed residues in the regulation of open complex formation and promoter DNA binding to be better understood. Their properties support the view that the interface between sigma54 and core RNAP is functionally specialised.
Figures
Similar articles
-
Interactions of regulated and deregulated forms of the sigma54 holoenzyme with heteroduplex promoter DNA.Nucleic Acids Res. 2002 Feb 15;30(4):886-93. doi: 10.1093/nar/30.4.886. Nucleic Acids Res. 2002. PMID: 11842099 Free PMC article.
-
Beta subunit residues 186-433 and 436-445 are commonly used by Esigma54 and Esigma70 RNA polymerase for open promoter complex formation.J Mol Biol. 2002 Jun 21;319(5):1067-83. doi: 10.1016/S0022-2836(02)00330-3. J Mol Biol. 2002. PMID: 12079348
-
The sigma 54 DNA-binding domain includes a determinant of enhancer responsiveness.Mol Microbiol. 1999 Sep;33(6):1200-9. doi: 10.1046/j.1365-2958.1999.01566.x. Mol Microbiol. 1999. PMID: 10510234
-
Transcriptional regulation under pressure conditions by RNA polymerase sigma54 factor with a two-component regulatory system in Shewanella violacea.Extremophiles. 2002 Apr;6(2):89-95. doi: 10.1007/s00792-001-0247-2. Extremophiles. 2002. PMID: 12013439 Review.
-
The bacterial enhancer-dependent RNA polymerase.Biochem J. 2016 Nov 1;473(21):3741-3753. doi: 10.1042/BCJ20160741C. Biochem J. 2016. PMID: 27789741 Free PMC article. Review.
Cited by
-
The role of bacterial enhancer binding proteins as specialized activators of σ54-dependent transcription.Microbiol Mol Biol Rev. 2012 Sep;76(3):497-529. doi: 10.1128/MMBR.00006-12. Microbiol Mol Biol Rev. 2012. PMID: 22933558 Free PMC article. Review.
-
Large-scale genetic analysis and biological traits of two SigB factors in Listeria monocytogenes: lineage correlations and differential functions.Front Microbiol. 2023 Nov 13;14:1268709. doi: 10.3389/fmicb.2023.1268709. eCollection 2023. Front Microbiol. 2023. PMID: 38029172 Free PMC article.
-
Regulated communication between the upstream face of RNA polymerase and the beta' subunit jaw domain.EMBO J. 2004 Oct 27;23(21):4264-74. doi: 10.1038/sj.emboj.7600407. Epub 2004 Oct 7. EMBO J. 2004. PMID: 15470503 Free PMC article.
-
Regulation of type VI secretion gene clusters by sigma54 and cognate enhancer binding proteins.J Bacteriol. 2011 May;193(9):2158-67. doi: 10.1128/JB.00029-11. Epub 2011 Mar 4. J Bacteriol. 2011. PMID: 21378190 Free PMC article.
-
Sigma54-dependent transcription activator phage shock protein F of Escherichia coli: a fragmentation approach to identify sequences that contribute to self-association.Biochem J. 2004 Mar 15;378(Pt 3):735-44. doi: 10.1042/BJ20031464. Biochem J. 2004. PMID: 14659000 Free PMC article.
References
-
- Wedel A. and Kustu,S. (1995) The bacterial enhancer-binding protein NtrC is a molecular machine: ATP hydrolysis is coupled to transcriptional activation. Genes Dev., 9, 2042–2052. - PubMed
-
- Popham D.L., Szeto,D., Keener,J. and Kustu,S. (1989) Purification of the alternative sigma factor, sigma 54, from Salmonella typhimurium and characterisation of sigma 54-holoenzyme. Science, 243, 629–635. - PubMed
-
- Wang L. and Gralla,J.D. (2001) Roles for the C-terminal region of sigma 54 in transcriptional silencing and DNA-binding. J. Biol. Chem., 276, 8979–8986. - PubMed
-
- Chaney M. and Buck,M. (1999) The sigma 54 DNA binding domain includes a determinant of enhancer responsiveness. Mol. Microbiol., 33, 1200–1209. - PubMed
