

Oxygen deficiency responsive gene expression in Chlamydomonas reinhardtii through a copper-sensing signal transduction pathway
- PMID: 11842150
- PMCID: PMC148909
- DOI: 10.1104/pp.010694
Oxygen deficiency responsive gene expression in Chlamydomonas reinhardtii through a copper-sensing signal transduction pathway
Abstract
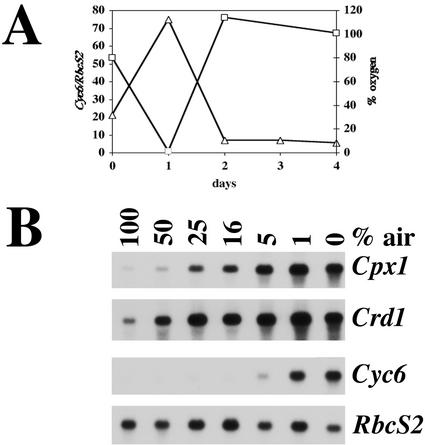
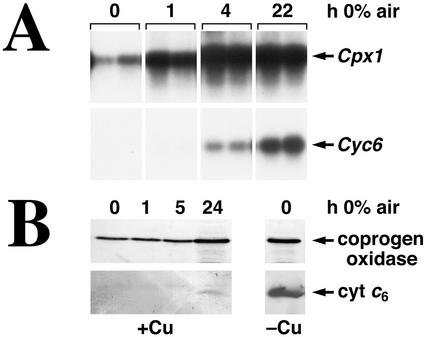
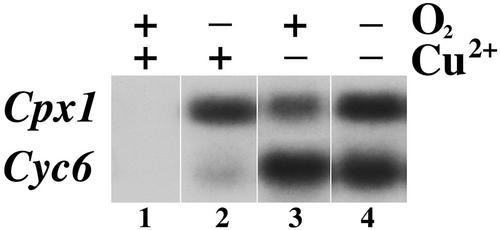
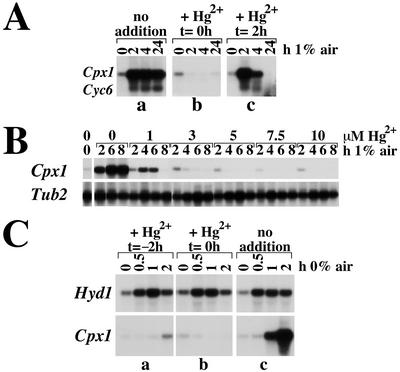
Chlamydomonas reinhardtii activates Cpx1, Cyc6, and Crd1, encoding, respectively, coproporphyrinogen oxidase, cytochrome c(6), and a novel di-iron enzyme when transferred to oxygen-deficient growth conditions. This response is physiologically relevant because C. reinhardtii experiences these growth conditions routinely, and furthermore, one of the target genes, Crd1, is functionally required for normal growth under oxygen-depleted conditions. The same genes are activated also in response to copper-deficiency through copper-response elements that function as target sites for a transcriptional activator. The core of the copper-response element, GTAC, is required also for the hypoxic response, as is a trans-acting locus, CRR1. Mercuric ions, which antagonize the copper-deficiency response, also antagonize the oxygen-deficiency response of these target genes. Taken together, these observations suggest that the oxygen- and copper-deficiency responses share signal transduction components. Nevertheless, whereas the copper-response element is sufficient for the nutritional copper response, the oxygen-deficiency response requires, in addition, a second cis-element, indicating that the response to oxygen depletion is not identical to the nutritional copper response. The distinction between the two responses is also supported by comparative analysis of the response of the target genes, Cyc6, Cpx1, and Crd1, to copper versus oxygen deficiency. A Crr1-independent pathway for Hyd1 expression in oxygen-depleted C. reinhardtii demonstrates the existence of multiple oxygen/redox-responsive circuits in this model organism.
Figures




References
-
- Bauer CE, Elsen S, Bird TH. Mechanisms for redox control of gene expression. Annu Rev Microbiol. 1999;53:495–523. - PubMed
-
- de Hostos EL, Schilling J, Grossman AR. Structure and expression of the gene encoding the periplasmic arylsulfatase of Chlamydomonas reinhardtii. Mol Gen Genet. 1989;218:229–239. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
