

Abscisic acid determines basal susceptibility of tomato to Botrytis cinerea and suppresses salicylic acid-dependent signaling mechanisms
- PMID: 11842153
- PMCID: PMC148912
- DOI: 10.1104/pp.010605
Abscisic acid determines basal susceptibility of tomato to Botrytis cinerea and suppresses salicylic acid-dependent signaling mechanisms
Abstract
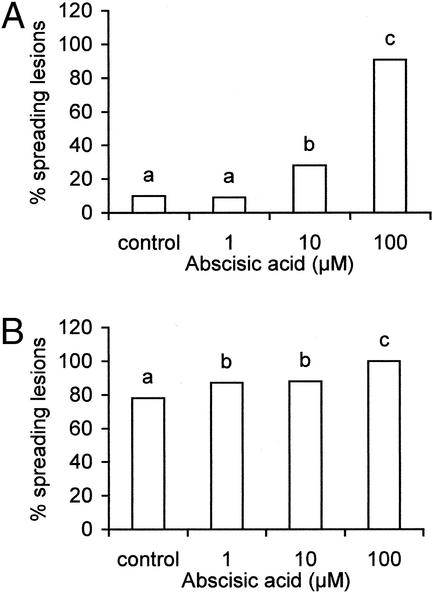
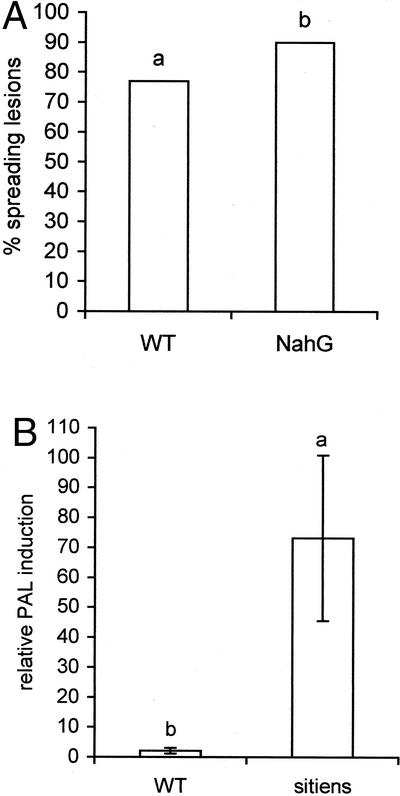
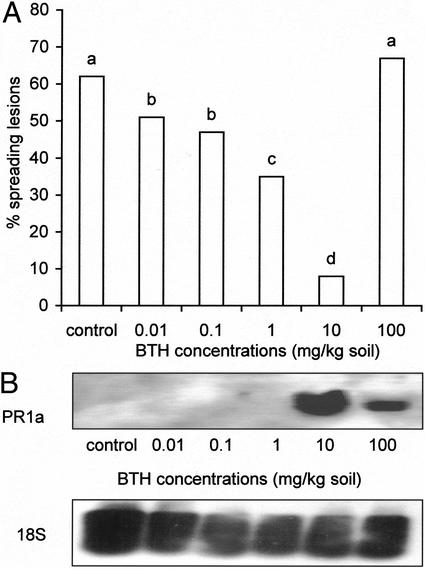
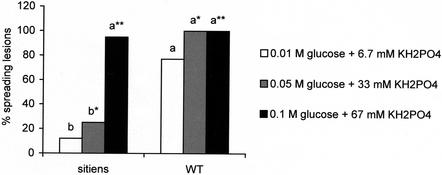
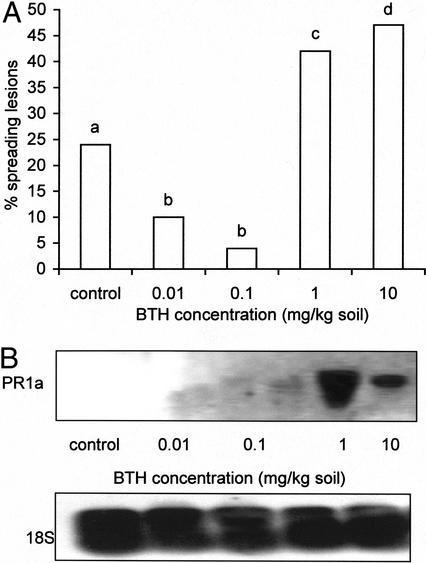
Abscisic acid (ABA) is one of the plant hormones involved in the interaction between plants and pathogens. In this work, we show that tomato (Lycopersicon esculentum Mill. cv Moneymaker) mutants with reduced ABA levels (sitiens plants) are much more resistant to the necrotrophic fungus Botrytis cinerea than wild-type (WT) plants. Exogenous application of ABA restored susceptibility to B. cinerea in sitiens plants and increased susceptibility in WT plants. These results indicate that ABA plays a major role in the susceptibility of tomato to B. cinerea. ABA appeared to interact with a functional plant defense response against B. cinerea. Experiments with transgenic NahG tomato plants and benzo(1,2,3)thiadiazole-7-carbothioic acid demonstrated the importance of salicylic acid in the tomato-B. cinerea interaction. In addition, upon infection with B. cinerea, sitiens plants showed a clear increase in phenylalanine ammonia lyase activity, which was not observed in infected WT plants, indicating that the ABA levels in healthy WT tomato plants partly repress phenylalanine ammonia lyase activity. In addition, sitiens plants became more sensitive to benzo(1,2,3)thiadiazole-7-carbothioic acid root treatment. The threshold values for PR1a gene expression declined with a factor 10 to 100 in sitiens compared with WT plants. Thus, ABA appears to negatively modulate the salicylic acid-dependent defense pathway in tomato, which may be one of the mechanisms by which ABA levels determine susceptibility to B. cinerea.
Figures




References
-
- Ausubel FM, Brent R, Kingston ER. Current Protocols in Molecular Biology. Brooklyn, NY: Current Protocols; 1993.
-
- Bothe H, Klingner A, Kaldorf M, Schmitz O, Esch H, Hundeshagen B, Kernebeck H. Biochemical approaches to the study of plant-fungal interactions in arbuscular mycorrhiza. Experientia. 1994;50:919–925.
-
- Brading PA, Hammond-Kosack KE, Parr A, Jones JDG. Salicylic acid is not required for Cf-2 and Cf-9-dependent resistance of tomato to Cladosporium fulvum. Plant J. 2000;23:305–318. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
