

Biosynthesis and distribution of chlorophyll among the photosystems during recovery of the green alga Dunaliella salina from irradiance stress
- PMID: 11842163
- PMCID: PMC148922
- DOI: 10.1104/pp.010595
Biosynthesis and distribution of chlorophyll among the photosystems during recovery of the green alga Dunaliella salina from irradiance stress
Abstract
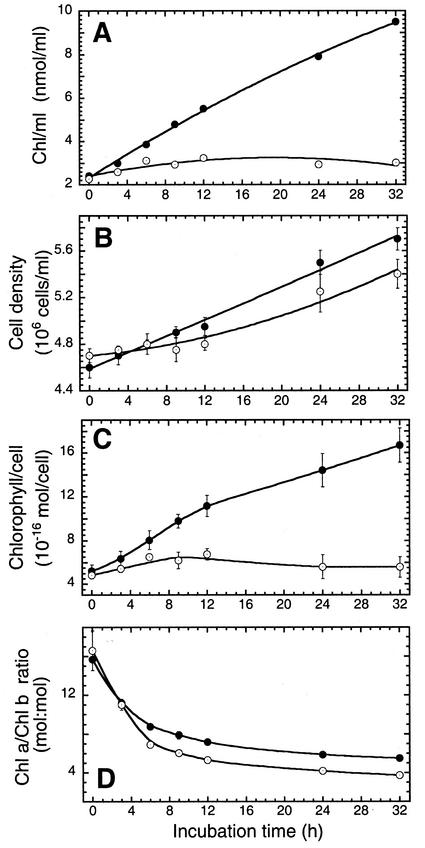
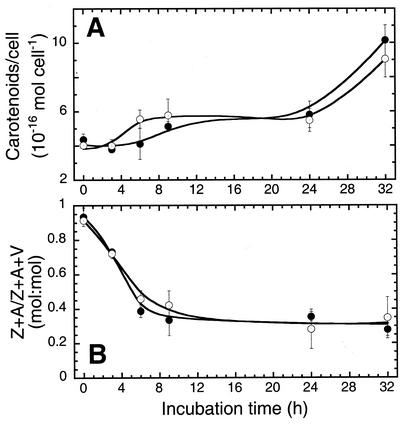
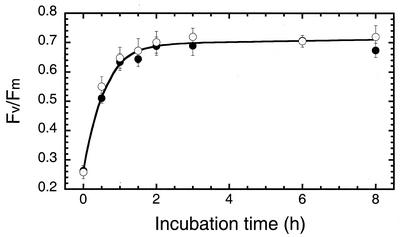
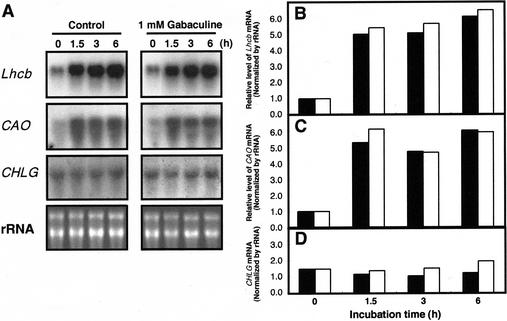
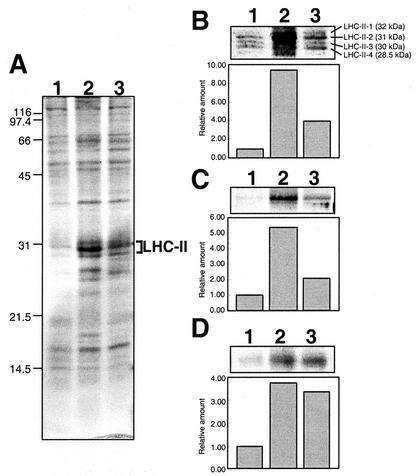
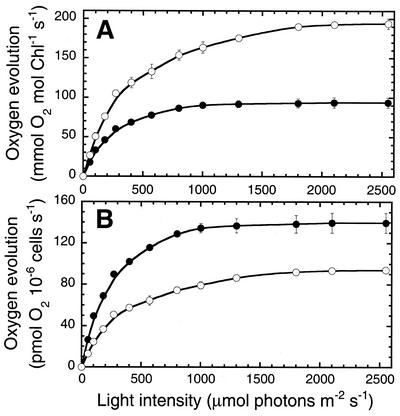
To elucidate the mechanism of an irradiance-dependent adjustment in the chlorophyll (Chl) antenna size of Dunaliella salina, we investigated the regulation of expression of the Chl a oxygenase (CAO) and light-harvesting complex b (Lhcb) genes as a function of Chl availability in the photosynthetic apparatus. After a high-light to low-light shift of the cultures, levels of both CAO and Lhcb transcripts were rapidly induced by about 6-fold and reached a high steady-state level within 1.5 h of the shift. This was accompanied by repair of photodamaged photosystem II (PSII) reaction centers, accumulation of Chl a and Chl b (4:1 ratio), photosystem I (PSI), light-harvesting complex, and by enlargement of the Chl antenna size of both photosystems. In gabaculine-treated cells, induction of CAO and Lhcb transcripts was not affected despite substantial inhibition in de novo Chl biosynthesis. However, cells were able to synthesize and accumulate some Chl a and Chl b (1:1 ratio), resulting in a marked lowering of the Chl a to Chl b ratio in the presence of this inhibitor. Assembly incorporation of light-harvesting complex and a corresponding Chl antenna size increase, mostly for the existing photosystems, was noted in the presence of gabaculine. Repair of photodamaged PSII was not affected by gabaculine. However, assembly accumulation of new PSI was limited under such conditions. These results suggest a coordinate regulation of CAO and Lhcb gene transcription by irradiance, independent of Chl availability. The results are discussed in terms of different signal transduction pathways for the regulation of the photosynthetic apparatus organization by irradiance.
Figures
References
-
- Akoyunoglou G, Argyroudi-Akoyunoglou J. Post-translation regulation of chloroplast differentiation. In: Akoyunoglou G, Senger H, editors. Regulation of chloroplast differentiation. A.R. New York: Liss; 1986. pp. 571–582.
-
- Anderson JM. Photoregulation of the composition, function and structure of thylakoid membranes. Annu Rev Plant Physiol. 1986;37:93–136.
-
- Aro E-M, Virgin I, Andersson B. Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim Biophys Acta. 1993;1143:113–134. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
