

Review
doi: 10.1128/JB.184.5.1225-1233.2002.
Mechanism of coupling of transport to hydrolysis in bacterial ATP-binding cassette transporters
Affiliations
- PMID: 11844750
- PMCID: PMC134847
- DOI: 10.1128/JB.184.5.1225-1233.2002
Item in Clipboard
Review
Mechanism of coupling of transport to hydrolysis in bacterial ATP-binding cassette transporters
J Bacteriol.
2002 Mar.
No abstract available
Figures
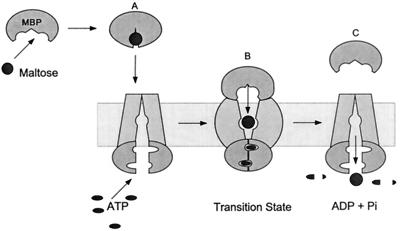
Model for maltose transport. (A) MBP binds maltose, undergoing a change from an open conformation to a closed conformation, generating a high-affinity sugar-binding site. In the closed conformation, MBP binds MalFGK2 to initiate transport and hydrolysis. (B) In the transition state for ATP hydrolysis, MBP becomes tightly bound to MalFGK2, and internal sugar-binding sites are exposed to each other. This opening of MBP in the transition state reduces the affinity of MBP for maltose, facilitating the transfer of sugar to MalFGK2. (C) Maltose is transported, and MBP is released after reexposure of the membrane-binding site to the cytoplasm. MBP activates the ATPase activity of MalK by bringing the two MalK subunits into close proximity, completing the nucleotide-binding site(s) at the MalK-MalK interface with residues donated from the opposing subunit. Reprinted from reference .
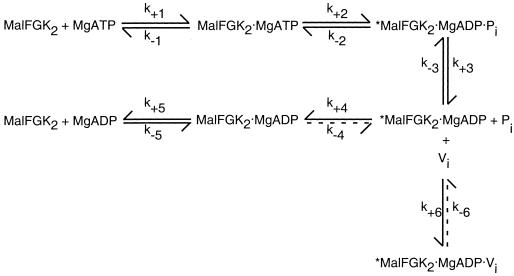
Scheme for ATP hydrolysis and vanadate inhibition. In this scheme for ATP hydrolysis by MalFGK2, ATP binding (step 1) and ATP hydrolysis (step 2) are followed by the ordered release of Pi (step 3) and then ADP (steps 4 and 5). Release of ADP is predicted to be a two-step process in which the transport complex undergoes a conformational change that is not readily reversible, from a high-energy species (∗) to a low-energy species (step 4) prior to ADP release (step 5). Vanadate will inhibit ATPase activity (step 6) only if it binds to the high-energy species formed immediately following ATP hydrolysis and Pi release (steps 1 to 3). Once formed, the vanadate-bound species is quite stable in the maltose system (107). Reprinted from reference with permission of the publisher.
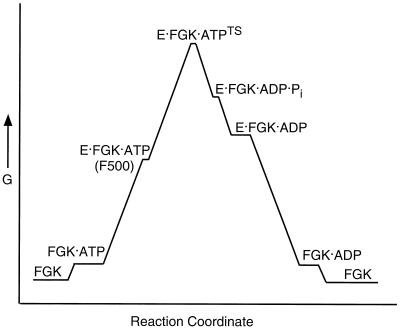
Free energy changes for MalFGK2 during ATP hydrolysis. For illustrative purposes, we have constructed this hypothetical energy diagram (G, Gibbs free energy) for MalFGK2 (FGK) during ATP hydrolysis. Binding of both ATP and MBP (E) to FGK will trigger ATP hydrolysis. E becomes more tightly bound to FGK as the complex approaches the transition state (EFGKTS), passing through an intermediate conformation that may resemble the binding protein-independent MalF500 mutant (63, 121). Following ATP hydrolysis, dissociation of Pi is likely to precede dissociation of ADP, allowing vanadate to bind and stabilize the complex in the transition state (TS) conformation. The ADP-bound intermediate may exist in both a high-energy and low-energy conformation, since ADP plus vanadate cannot trap the transition state.
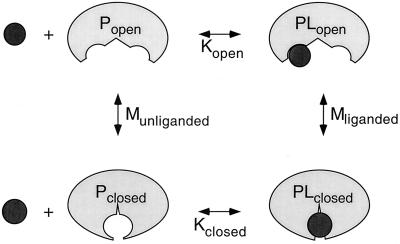
Thermodynamics of ligand and ligand-binding protein interactions. Munliganded and Mliganded are the equilibrium allosteric constants for the interconversion of the open and closed forms of the binding protein in the absence and presence of ligand (L), respectively. Kopen and Kclosed are the equilibrium association constants for the binding of ligand to the open and closed forms of the binding protein, respectively (see equations 1 to 4). 



References
-
- Aleksandrov, L., A. Mengos, X. Chang, A. Aleksandrov, and J. R. Riordan. 2001. Differential interactions of nucleotides at the two nucleotide binding domains of the cystic fibrosis transmembrane conductance regulator. J. Biol. Chem. 276:12918-12923. - PubMed
-
- Al-Shawi, M. K., and A. E. Senior. 1993. Characterization of the adenosine triphosphatase activity of Chinese hamster P-glycoprotein. J. Biol. Chem. 268:4197-4206. - PubMed
-
- Al-Shawi, M. K., I. L. Urbatsch, and A. E. Senior. 1994. Covalent inhibitors of P-glycoprotein ATPase activity. J. Biol. Chem. 269:8986-8992. - PubMed
-
- Ames, G. F., C. S. Mimura, and V. Shyamala. 1990. Bacterial periplasmic permeases belong to a family of transport proteins operating from Escherichia coli to humans: traffic ATPases. FEMS Microbiol. Rev. 6:429-446. - PubMed
-
- Ames, G. F. L., C. E. Liu, A. K. Joshi, and K. Nikaido. 1996. Liganded and unliganded receptors interact with equal affinity with the membrane complex of periplasmic permeases, a subfamily of traffic ATPases. J. Biol. Chem. 271:14264-14270. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
