

Hippocampal population activity during the small-amplitude irregular activity state in the rat
- PMID: 11850464
- PMCID: PMC6757571
- DOI: 10.1523/JNEUROSCI.22-04-01373.2002
Hippocampal population activity during the small-amplitude irregular activity state in the rat
Abstract
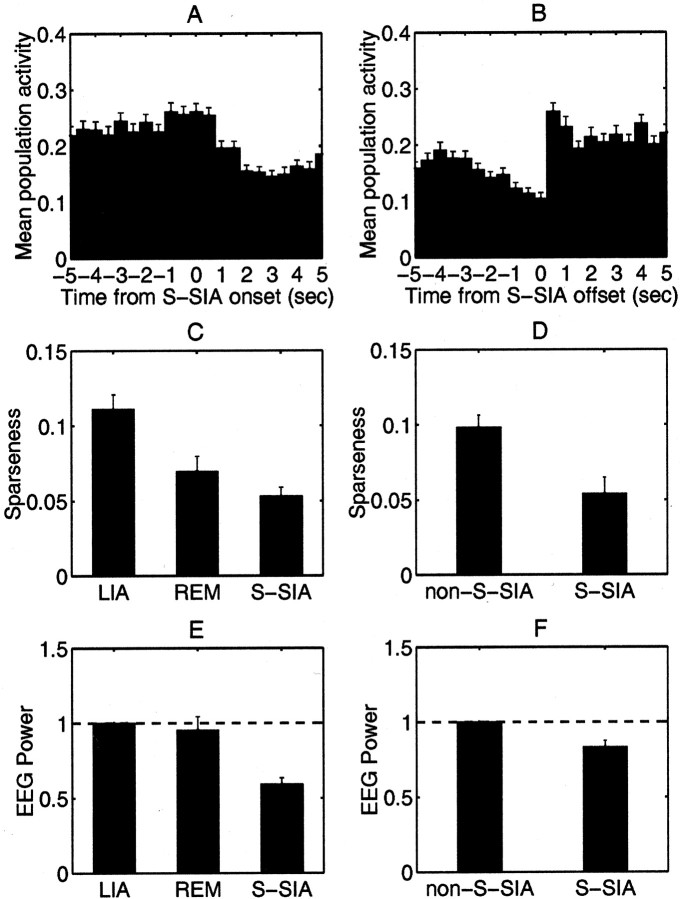
The sleeping rat cycles between two well-characterized physiological states, slow-wave sleep (SWS) and rapid-eye-movement sleep (REM), often identified by the presence of large-amplitude irregular activity (LIA) and theta activity, respectively, in the hippocampal EEG. Inspection of the activity of ensembles of hippocampal CA1 complex-spike cells along with the EEG reveals the presence of a third physiological state within SWS. We characterize the hippocampal EEG and population activity of this third state relative to theta activity and LIA, its incidence relative to REM and LIA, and the functional correlates of its population activity. This state occurs repeatedly within stretches of SWS, occupying approximately 33% of SWS and approximately 20% of total sleep, and it follows nearly every REM episode; however, it never occurs just before a REM episode. The EEG during this state becomes low in amplitude for a few seconds, probably corresponding to "small-amplitude irregular activity" (SIA) described in the literature; we will call its manifestation during sleep "S-SIA." During S-SIA, a small subset of cells becomes active, whereas the rest remain nearly silent, with the same subset of cells active across long sequences of S-SIA episodes. These cells are physiologically indistinguishable from ordinary complex-spike cells; thus, the question arises as to whether they have any special functional correlates. Indeed, many of these cells are found to have place fields encompassing the location where the rat sleeps, raising the possibility that S-SIA is a state of increased alertness in which the animal's location in the environment is represented in the brain.
Figures






Similar articles
-
Hippocampal place cells are not controlled by visual input during the small irregular activity state in the rat.J Neurosci. 2004 May 26;24(21):5070-7. doi: 10.1523/JNEUROSCI.5650-03.2004. J Neurosci. 2004. PMID: 15163700 Free PMC article.
-
Level of arousal during the small irregular activity state in the rat hippocampal EEG.J Neurophysiol. 2004 Jun;91(6):2649-57. doi: 10.1152/jn.01082.2003. Epub 2004 Jan 28. J Neurophysiol. 2004. PMID: 14749309
-
Brain stem gigantocellular neurons: patterns of activity during behavior and sleep in the freely moving rat.J Neurophysiol. 1979 Jan;42(1 Pt 1):214-28. doi: 10.1152/jn.1979.42.1.214. J Neurophysiol. 1979. PMID: 219157
-
The future of computer-assisted investigation of the polysomnogram: sleep microstructure.J Clin Neurophysiol. 1996 Jul;13(4):285-94. doi: 10.1097/00004691-199607000-00003. J Clin Neurophysiol. 1996. PMID: 8858491 Review.
-
The visual scoring of sleep and arousal in infants and children.J Clin Sleep Med. 2007 Mar 15;3(2):201-40. J Clin Sleep Med. 2007. PMID: 17557427 Review.
Cited by
-
Hippocampal place cells are not controlled by visual input during the small irregular activity state in the rat.J Neurosci. 2004 May 26;24(21):5070-7. doi: 10.1523/JNEUROSCI.5650-03.2004. J Neurosci. 2004. PMID: 15163700 Free PMC article.
-
Hippocampal theta in the newborn rat is revealed under conditions that promote REM sleep.J Neurosci. 2003 Feb 15;23(4):1114-8. doi: 10.1523/JNEUROSCI.23-04-01114.2003. J Neurosci. 2003. PMID: 12598598 Free PMC article.
-
Hippocampal and cortical communication around micro-arousals in slow-wave sleep.Sci Rep. 2019 Apr 10;9(1):5876. doi: 10.1038/s41598-019-42100-5. Sci Rep. 2019. PMID: 30971751 Free PMC article.
-
Stored-trace reactivation in rat prefrontal cortex is correlated with down-to-up state fluctuation density.J Neurosci. 2010 Feb 17;30(7):2650-61. doi: 10.1523/JNEUROSCI.1617-09.2010. J Neurosci. 2010. PMID: 20164349 Free PMC article.
-
A hippocampal network for spatial coding during immobility and sleep.Nature. 2016 Mar 10;531(7593):185-90. doi: 10.1038/nature17144. Epub 2016 Mar 2. Nature. 2016. PMID: 26934224 Free PMC article.
References
-
- Bergmann BM, Winter JB, Rosenberg RS, Rechtschaffen A. NREM sleep with low-voltage EEG in the rat. Sleep. 1987;10:1–11. - PubMed
-
- Buzsáki G. Hippocampal sharp waves: their origin and significance. Brain Res. 1986;298:242–252. - PubMed
-
- Buzsáki G, Horvath Z, Urioste R, Hetke J, Wise K. High-frequency network oscillation in the hippocampus. Science. 1992;256:1025–1027. - PubMed
-
- Dement W, Kleitman N. The relation of eye movements during sleep to dream activity: an objective method for the study of dreaming. J Exp Psychol. 1957a;53:339–346. - PubMed
-
- Dement W, Kleitman N. Cyclic variations in EEG during sleep and their relation to eye movements, body motility, and dreaming. Electroencephalogr Clin Neurophysiol. 1957b;9:673–690. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous