

Regulation of the glutamine transporter SN1 by extracellular pH and intracellular sodium ions
- PMID: 11850497
- PMCID: PMC2290136
- DOI: 10.1113/jphysiol.2001.013303
Regulation of the glutamine transporter SN1 by extracellular pH and intracellular sodium ions
Abstract
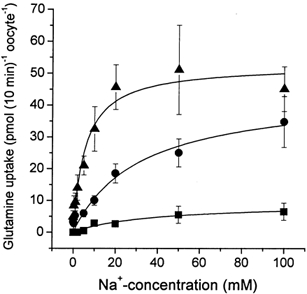
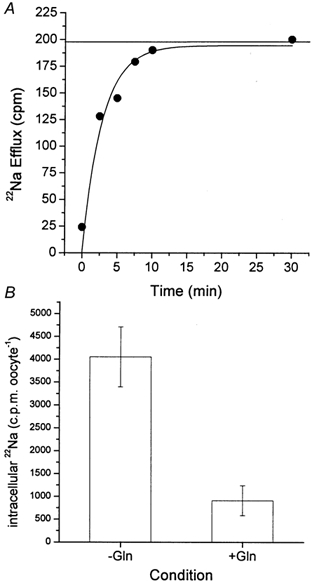
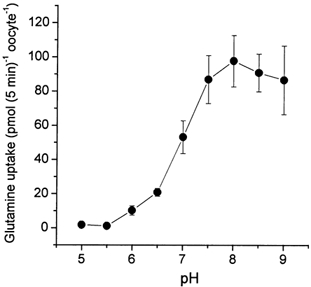
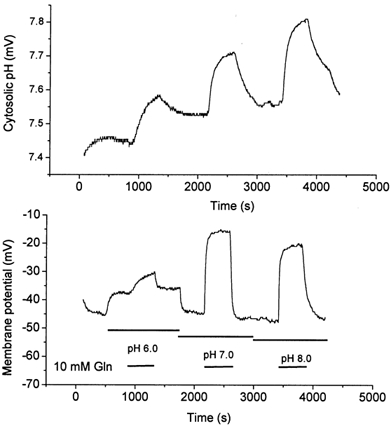
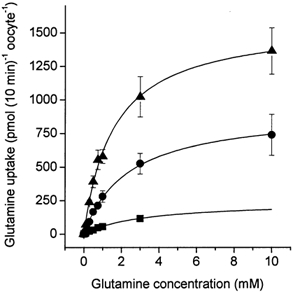
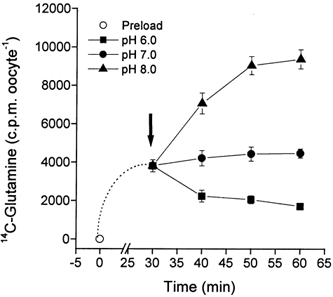
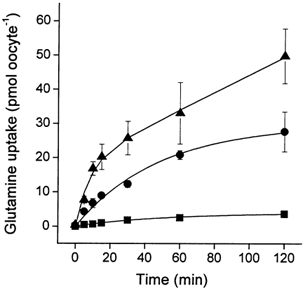
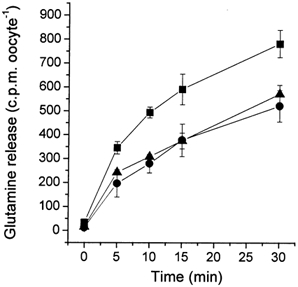
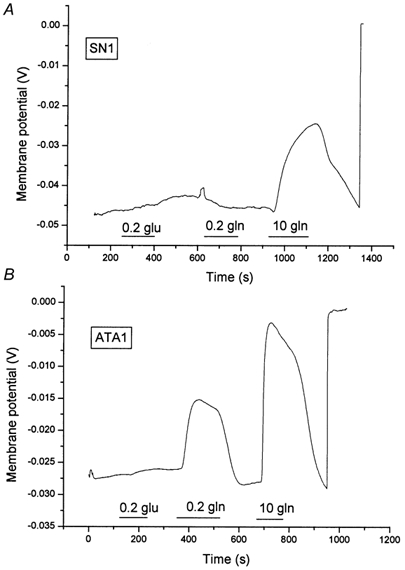
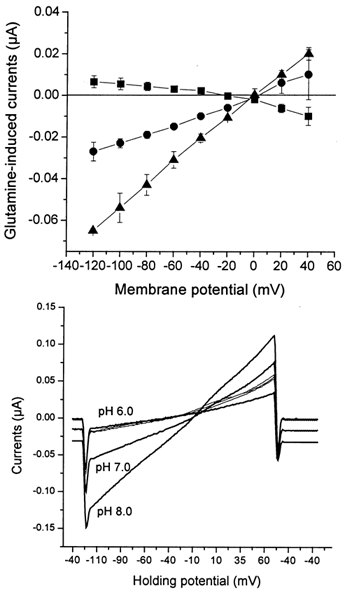
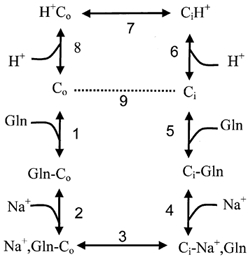
The glutamine transporter SN1 has recently been identified as one of the major glutamine transporters in hepatocytes and brain astrocytes. It appears to be the molecular correlate of system N amino acid transport. Two different transport mechanisms have been proposed for this transporter. These are an electroneutral mechanism, in which glutamine uptake is coupled to an exchange of 1Na+ and 1H+, or an electrogenic mechanism coupled to the exchange of 2Na+ against 1H+. This study was performed to solve these discrepancies and to investigate the reversibility of the transporter. When SN1 was expressed in Xenopus laevis oocytes, glutamine uptake was accompanied by a cotransport of 2-3 Na+ ions as determined by 22Na+ fluxes. However, at the same time a rapid release of intracellular Na+ was observed indicating an active exchange of Na+ ions. The driving force of the proton electrochemical gradient was equivalent to that of the sodium electrochemical gradient. Acidification of the extracellular medium caused the transporter to run in reverse and to release glutamine. Determination of accumulation ratios at different driving forces were in agreement with an electroneutral 1Na+-glutamine cotransport-1H+ antiport. Inward currents that were observed during glutamine uptake were much smaller than expected for a stoichiometric cotransport of charges. A slippage mode in the transporter mechanism and pH-regulated endogenous oocyte cation channels are likely to contribute to the observed currents.
Figures
Similar articles
-
Primary structure, genomic organization, and functional and electrogenic characteristics of human system N 1, a Na+- and H+-coupled glutamine transporter.J Biol Chem. 2000 Aug 4;275(31):23707-17. doi: 10.1074/jbc.M002282200. J Biol Chem. 2000. PMID: 10823827
-
Heterologous expression of the glutamine transporter SNAT3 in Xenopus oocytes is associated with four modes of uncoupled transport.J Biol Chem. 2007 Feb 9;282(6):3788-98. doi: 10.1074/jbc.M609452200. Epub 2006 Dec 5. J Biol Chem. 2007. PMID: 17148440
-
Glutamine efflux from astrocytes is mediated by multiple pathways.J Neurochem. 2003 Oct;87(1):127-35. doi: 10.1046/j.1471-4159.2003.01981.x. J Neurochem. 2003. PMID: 12969260
-
Molecular characteristics of mammalian and insect amino acid transporters: implications for amino acid homeostasis.J Exp Biol. 1997 Jan;200(Pt 2):269-86. doi: 10.1242/jeb.200.2.269. J Exp Biol. 1997. PMID: 9050235 Review.
-
Biology of the 2Na+/1H+ antiporter in invertebrates.J Exp Zool. 2001 Apr 1;289(4):232-44. doi: 10.1002/1097-010x(20010401/30)289:4<232::aid-jez4>3.0.co;2-t. J Exp Zool. 2001. PMID: 11241394 Review.
Cited by
-
Impact of Inhibition of Glutamine and Alanine Transport on Cerebellar Glial and Neuronal Metabolism.Biomolecules. 2022 Aug 27;12(9):1189. doi: 10.3390/biom12091189. Biomolecules. 2022. PMID: 36139028 Free PMC article.
-
Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release.Cells. 2019 Feb 20;8(2):184. doi: 10.3390/cells8020184. Cells. 2019. PMID: 30791579 Free PMC article. Review.
-
In vivo N-15 MRS study of glutamate metabolism in the rat brain.Anal Biochem. 2017 Jul 15;529:179-192. doi: 10.1016/j.ab.2016.08.025. Epub 2016 Aug 28. Anal Biochem. 2017. PMID: 27580850 Free PMC article. Review.
-
Sodium-Dependent Neutral Amino Acid Transporter 2 Can Serve as a Tertiary Carrier for l-Type Amino Acid Transporter 1-Utilizing Prodrugs.Mol Pharm. 2023 Feb 6;20(2):1331-1346. doi: 10.1021/acs.molpharmaceut.2c00948. Epub 2023 Jan 23. Mol Pharm. 2023. PMID: 36688491 Free PMC article.
-
Astrocyte-Neuron Interaction via the Glutamate-Glutamine Cycle and Its Dysfunction in Tau-Dependent Neurodegeneration.Int J Mol Sci. 2024 Mar 6;25(5):3050. doi: 10.3390/ijms25053050. Int J Mol Sci. 2024. PMID: 38474295 Free PMC article. Review.
References
-
- Albers A, Bröer A, Wagner CA, Setiawan I, Lang P, Kranz EU, Lang F, Bröer S. Na+ transport by the neural glutamine transporter ATA1. Pflügers Archiv. 2001;443:92–101. - PubMed
-
- Bröer S, Brookes N. Transfer of glutamine between astrocytes and neurons. Journal of Neurochemistry. 2001;77:705–719. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases