

Coordination of opposite-polarity microtubule motors
- PMID: 11854311
- PMCID: PMC2174082
- DOI: 10.1083/jcb.200109047
Coordination of opposite-polarity microtubule motors
Abstract
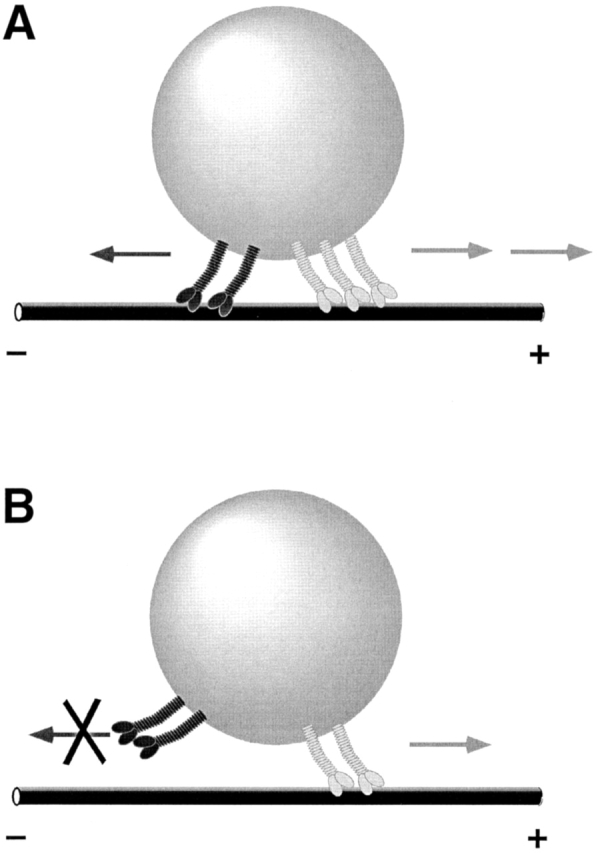
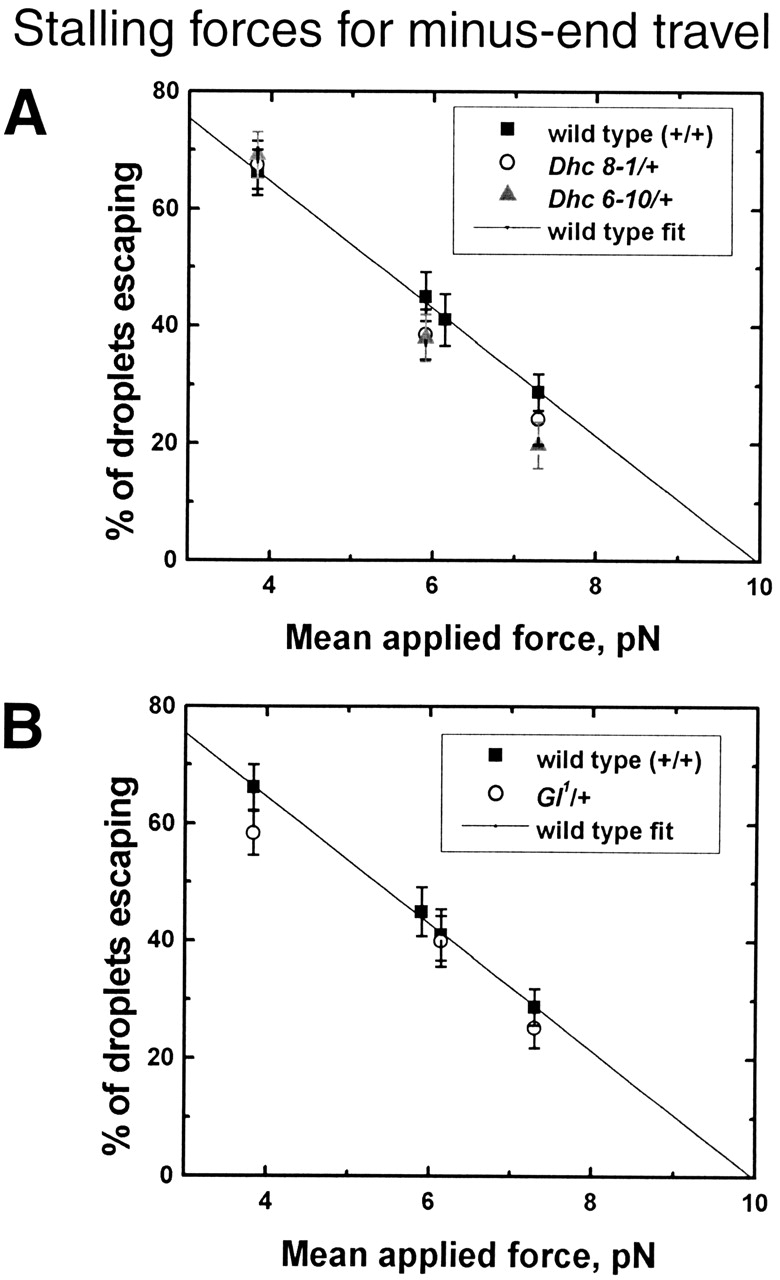
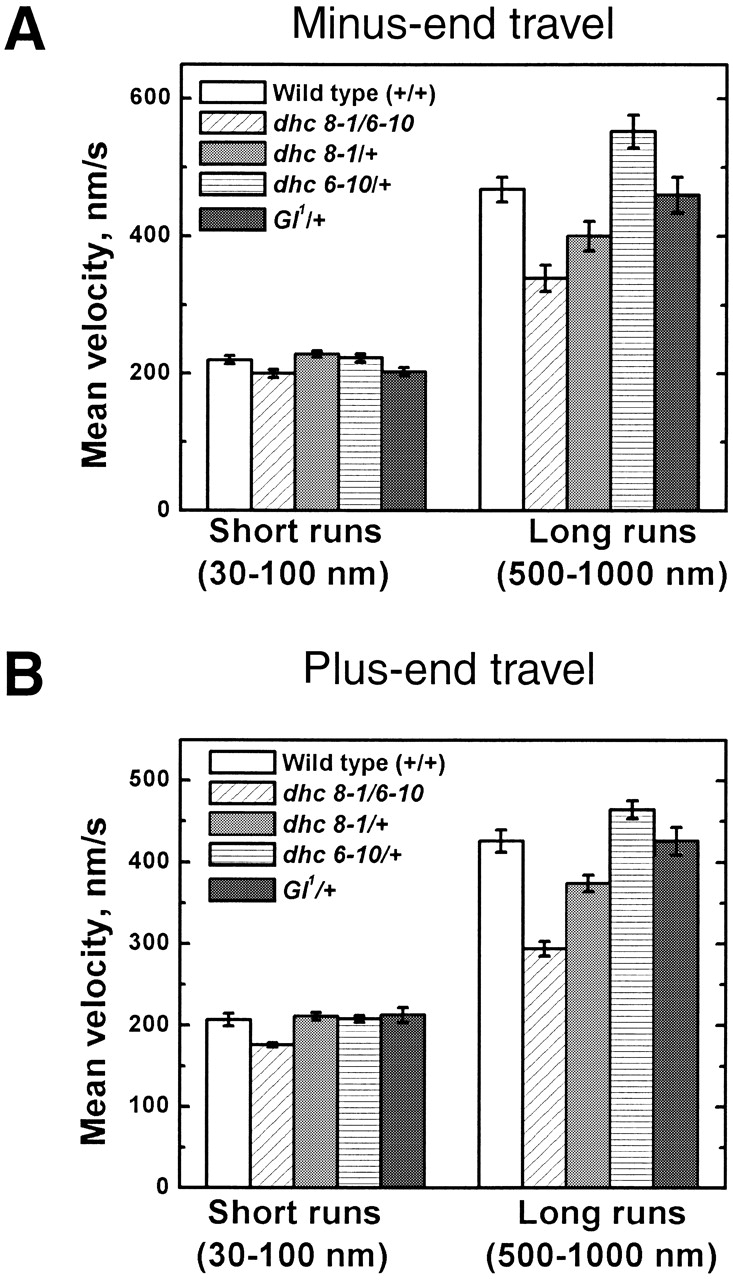
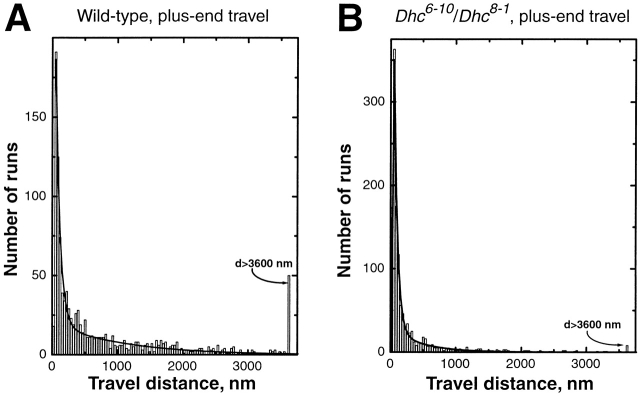
Many cargoes move bidirectionally, frequently reversing course between plus- and minus-end microtubule travel. For such cargoes, the extent and importance of interactions between the opposite-polarity motors is unknown. In this paper we test whether opposite-polarity motors on lipid droplets in Drosophila embryos are coordinated and avoid interfering with each other's activity, or whether they engage in a tug of war. To this end we impaired the minus-end transport machinery using dynein and dynactin mutations, and then investigated whether plus-end motion was improved or disrupted. We observe a surprisingly severe impairment of plus-end motion due to these alterations of minus-end motor activity. These observations are consistent with a coordination hypothesis, but cannot be easily explained with a tug of war model. Our measurements indicate that dynactin plays a crucial role in the coordination of plus- and minus-end-directed motors. Specifically, we propose that dynactin enables dynein to participate efficiently in bidirectional transport, increasing its ability to stay "on" during minus-end motion and keeping it "off" during plus-end motion.
Figures

Similar articles
-
Hook3 is a scaffold for the opposite-polarity microtubule-based motors cytoplasmic dynein-1 and KIF1C.J Cell Biol. 2019 Sep 2;218(9):2982-3001. doi: 10.1083/jcb.201812170. Epub 2019 Jul 18. J Cell Biol. 2019. PMID: 31320392 Free PMC article.
-
Kinesin-1, -2, and -3 motors use family-specific mechanochemical strategies to effectively compete with dynein during bidirectional transport.Elife. 2022 Sep 20;11:e82228. doi: 10.7554/eLife.82228. Elife. 2022. PMID: 36125250 Free PMC article.
-
Bidirectional transport of organelles: unity and struggle of opposing motors.Cell Biol Int. 2012 Jan;36(1):1-6. doi: 10.1042/CBI20110413. Cell Biol Int. 2012. PMID: 22142363 Review.
-
Cooperative Accumulation of Dynein-Dynactin at Microtubule Minus-Ends Drives Microtubule Network Reorganization.Dev Cell. 2018 Jan 22;44(2):233-247.e4. doi: 10.1016/j.devcel.2017.12.023. Dev Cell. 2018. PMID: 29401420 Free PMC article.
-
Bidirectional transport along microtubules.Curr Biol. 2004 Jul 13;14(13):R525-37. doi: 10.1016/j.cub.2004.06.045. Curr Biol. 2004. PMID: 15242636 Review.
Cited by
-
Motility Plays an Important Role in the Lifetime of Mammalian Lipid Droplets.Int J Mol Sci. 2021 Apr 7;22(8):3802. doi: 10.3390/ijms22083802. Int J Mol Sci. 2021. PMID: 33916886 Free PMC article. Review.
-
On the use of in vivo cargo velocity as a biophysical marker.Biochem Biophys Res Commun. 2007 Feb 16;353(3):835-40. doi: 10.1016/j.bbrc.2006.12.120. Epub 2006 Dec 22. Biochem Biophys Res Commun. 2007. PMID: 17196170 Free PMC article.
-
Axonal Transport: A Constrained System.J Neurol Neuromedicine. 2017;2(3):20-24. doi: 10.29245/2572.942X/2017/3.1118. Epub 2017 Mar 21. J Neurol Neuromedicine. 2017. PMID: 30467560 Free PMC article.
-
Dynactin is required for bidirectional organelle transport.J Cell Biol. 2003 Feb 3;160(3):297-301. doi: 10.1083/jcb.200210066. Epub 2003 Jan 27. J Cell Biol. 2003. PMID: 12551954 Free PMC article.
-
Live cell imaging reveals differential modifications to cytoplasmic dynein properties by phospho- and dephosphomimic mutations of the intermediate chain 2C S84.J Neurosci Res. 2014 Sep;92(9):1143-54. doi: 10.1002/jnr.23388. Epub 2014 May 5. J Neurosci Res. 2014. PMID: 24798412 Free PMC article.
References
-
- Blangy, A., L. Arnaud, and E.A. Nigg. 1997. Phosphorylation by p34cdc2 protein kinase regulates binding of the kinesin-related motor HsEg5 to the dynactin subunit p150. J. Biol. Chem. 272:19418–19424. - PubMed
-
- Fan, S.S., and D.F. Ready. 1997. Glued participates in distinct microtubule-based activities in Drosophila eye development. Development. 124:1497–1507. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
