

Pag1p, a novel protein associated with protein kinase Cbk1p, is required for cell morphogenesis and proliferation in Saccharomyces cerevisiae
- PMID: 11854408
- PMCID: PMC65645
- DOI: 10.1091/mbc.01-07-0365
Pag1p, a novel protein associated with protein kinase Cbk1p, is required for cell morphogenesis and proliferation in Saccharomyces cerevisiae
Abstract
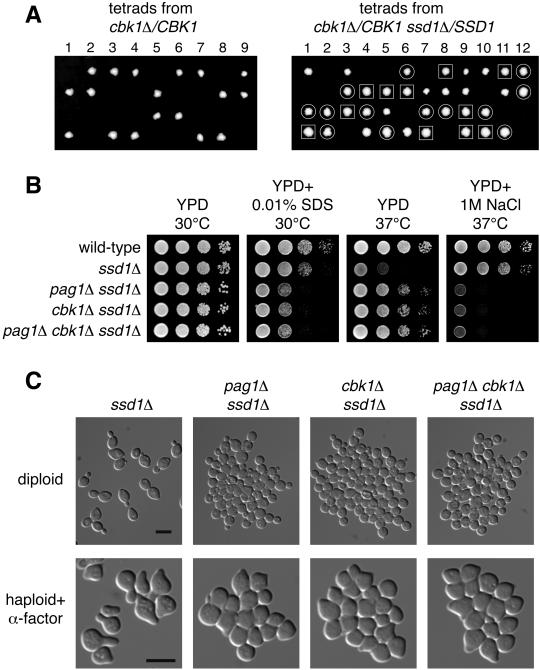
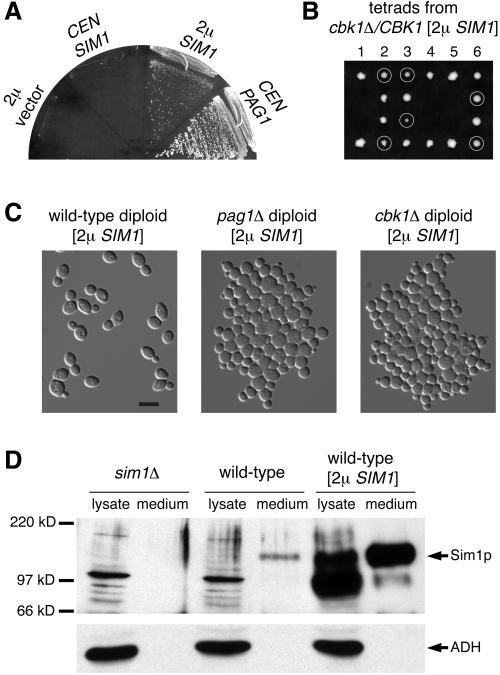
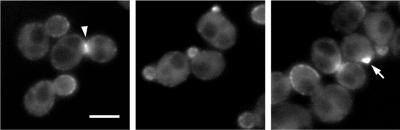
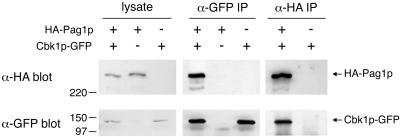
Protein kinases in the Cot-1/Orb6/Ndr/Warts family are important regulators of cell morphogenesis and proliferation. Cbk1p, a member of this family in Saccharomyces cerevisiae, has previously been shown to be required for normal morphogenesis in vegetatively growing cells and in haploid cells responding to mating pheromone. A mutant of PAG1, a novel gene in S. cerevisiae, displayed defects similar to those of cbk1 mutants. pag1 and cbk1 mutants share a common set of suppressors, including the disruption of SSD1, a gene encoding an RNA binding protein, and the overexpression of Sim1p, an extracellular protein. These genetic results suggest that PAG1 and CBK1 act in the same pathway. Furthermore, we found that Pag1p and Cbk1p localize to the same polarized peripheral sites and that they coimmunoprecipitate with each other. Pag1p is a conserved protein. The homologs of Pag1p in other organisms are likely to form complexes with the Cbk1p-related kinases and function with those kinases in the same biological processes.
Figures
References
-
- Adams A, Gottschling DE, Kaiser CA, Stearns T. Methods in Yeast Genetics. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory; 1997.
-
- Cong J, Geng W, He B, Liu J, Charlton J, Adler PN. The furry gene of Drosophila is important for maintaining the integrity of cellular extensions during morphogenesis. Development. 2001;128:2793–2802. - PubMed
-
- Cormack BP, Bertram G, Egerton M, Gow NA, Falkow S, Brown AJ. Yeast-enhanced green fluorescent protein (yEGFP) Microbiology. 1997;143:303–311. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
