

Mutation of YCS4, a budding yeast condensin subunit, affects mitotic and nonmitotic chromosome behavior
- PMID: 11854418
- PMCID: PMC65655
- DOI: 10.1091/mbc.01-05-0264
Mutation of YCS4, a budding yeast condensin subunit, affects mitotic and nonmitotic chromosome behavior
Abstract
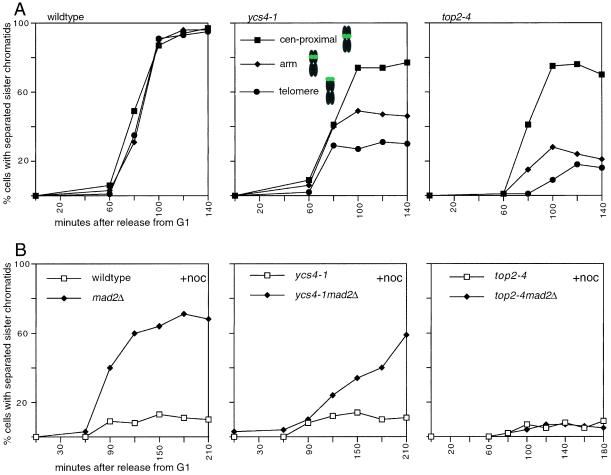
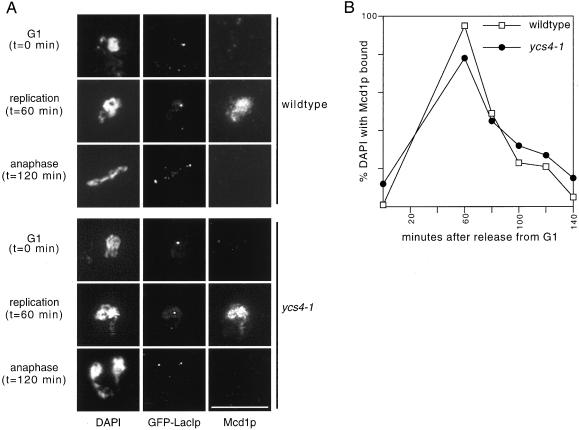
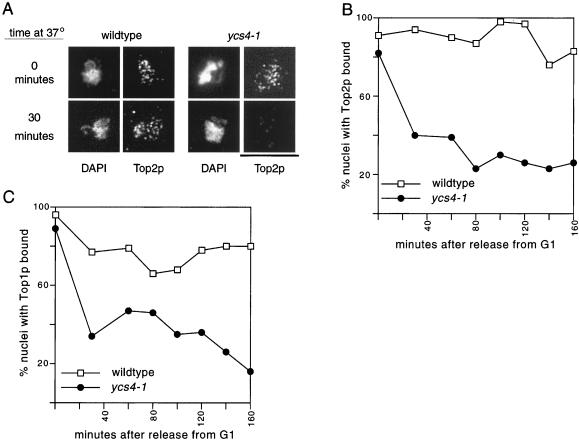
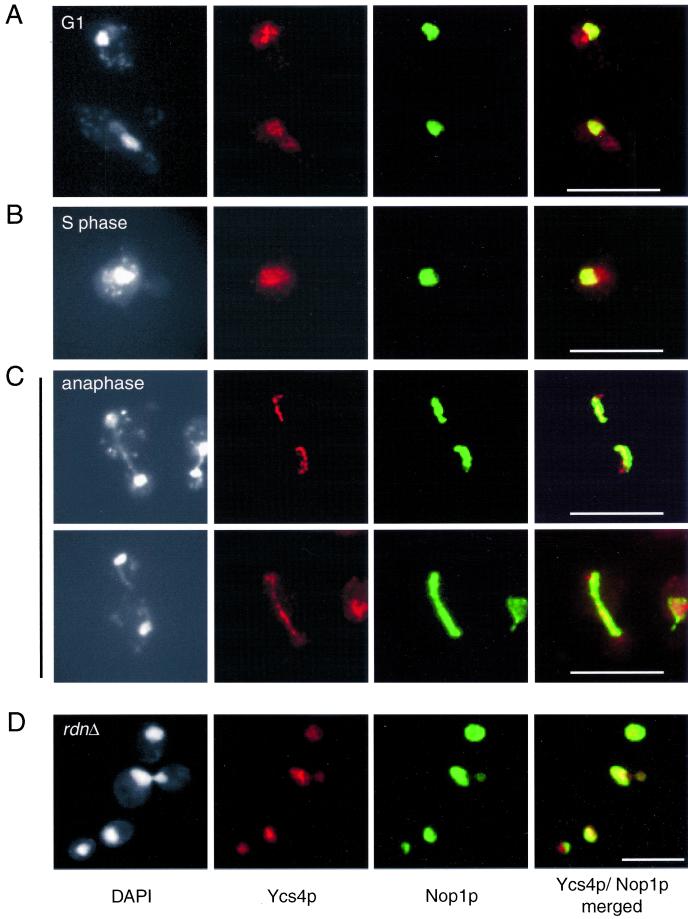
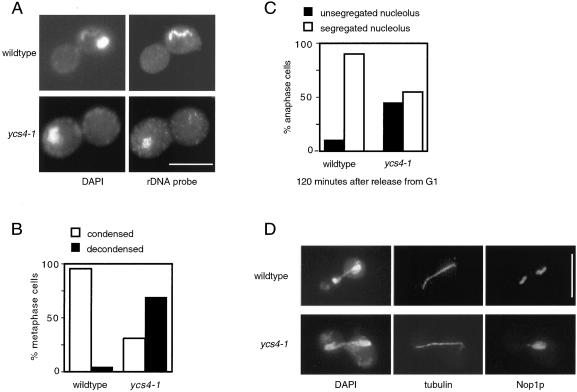
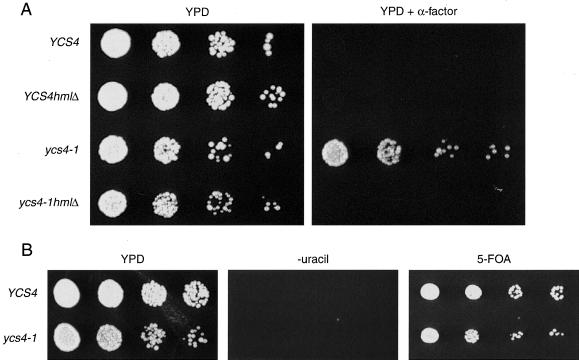
The budding yeast YCS4 gene encodes a conserved regulatory subunit of the condensin complex. We isolated an allele of this gene in a screen for mutants defective in sister chromatid separation or segregation. The phenotype of the ycs4-1 mutant is similar to topoisomerase II mutants and distinct from the esp1-1 mutant: the topological resolution of sister chromatids is compromised in ycs4-1 despite normal removal of cohesins from mitotic chromosomes. Consistent with a role in sister separation, YCS4 function is required to localize DNA topoisomerase I and II to chromosomes. Unlike its homologs in Xenopus and fission yeast, Ycs4p is associated with chromatin throughout the cell cycle; the only change in localization occurs during anaphase when the protein is enriched at the nucleolus. This relocalization may reveal the specific challenge that segregation of the transcriptionally hyperactive, repetitive array of rDNA genes can present during mitosis. Indeed, segregation of the nucleolus is abnormal in ycs4-1 at the nonpermissive temperature. Interrepeat recombination in the rDNA array is specifically elevated in ycs4-1 at the permissive temperature, suggesting that the Ycs4p plays a role at the array aside from its segregation. Furthermore, ycs4-1 is defective in silencing at the mating type loci at the permissive temperature. Taken together, our data suggest that there are mitotic as well as nonmitotic chromosomal abnormalities associated with loss of condensin function in budding yeast.
Figures
References
-
- Bhat MA, Philp AV, Glover DM, Bellen HJ. Chromatid segregation at anaphase requires the barren product, chromosome-associated protein that interacts with Topoisomerase II. Cell. 1996;87:1103–1114. - PubMed
-
- Biggins S, Murray AW. Sister chromatid cohesion in mitosis. Curr Opin Genet Dev. 1999;9:230–236. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
