

Repression of African swine fever virus polyprotein pp220-encoding gene leads to the assembly of icosahedral core-less particles
- PMID: 11861832
- PMCID: PMC135994
- DOI: 10.1128/jvi.76.6.2654-2666.2002
Repression of African swine fever virus polyprotein pp220-encoding gene leads to the assembly of icosahedral core-less particles
Abstract
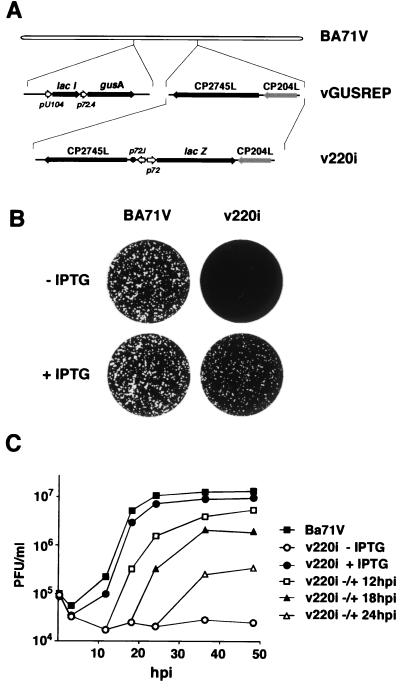
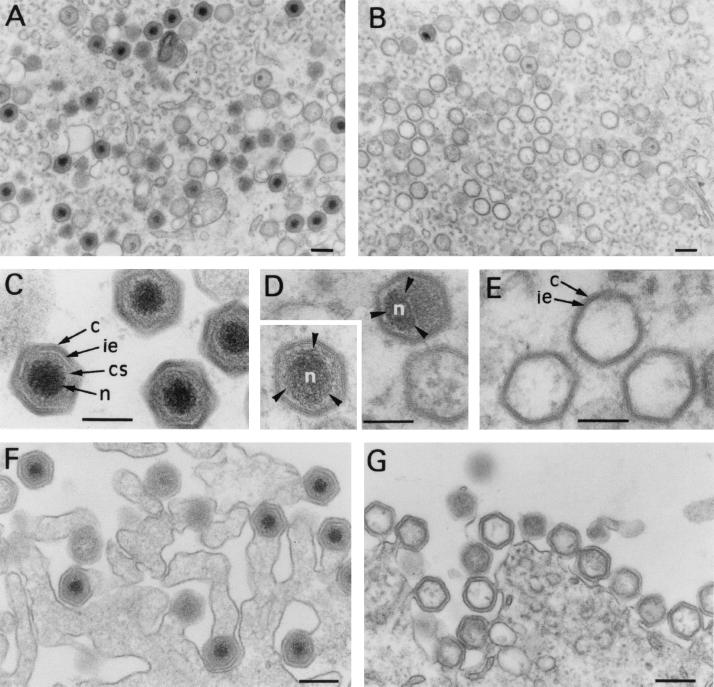
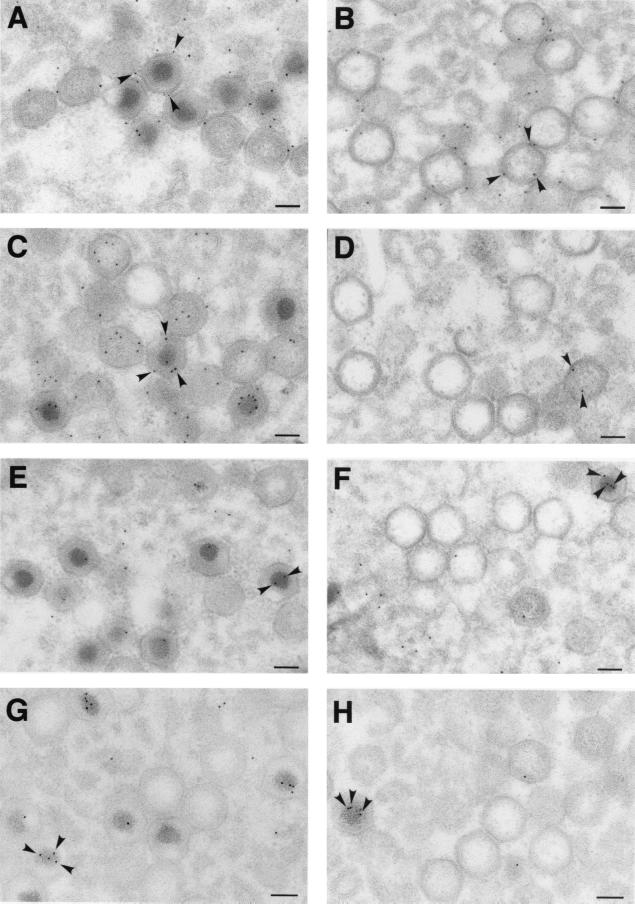
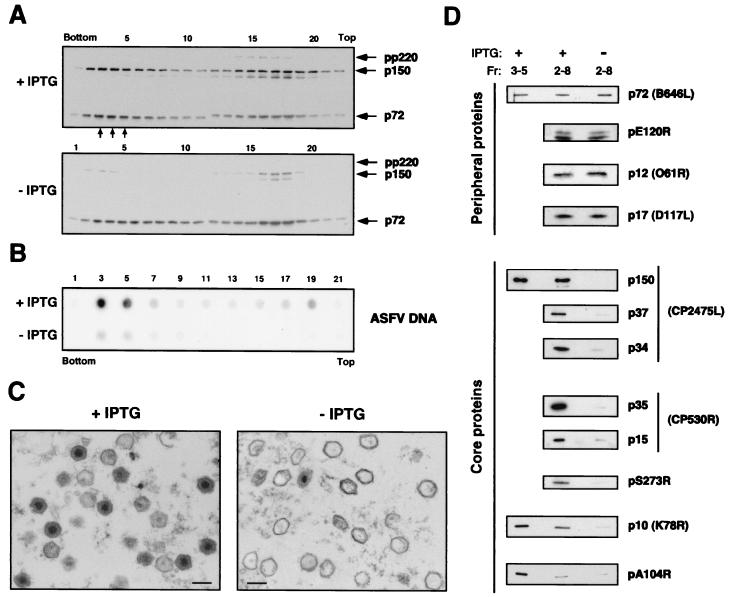
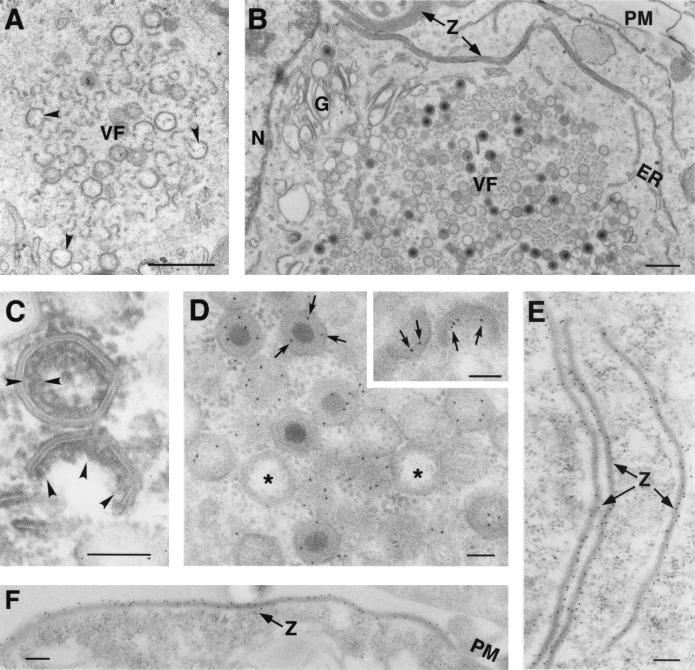
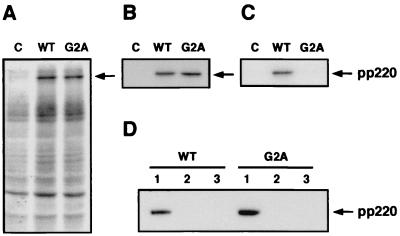
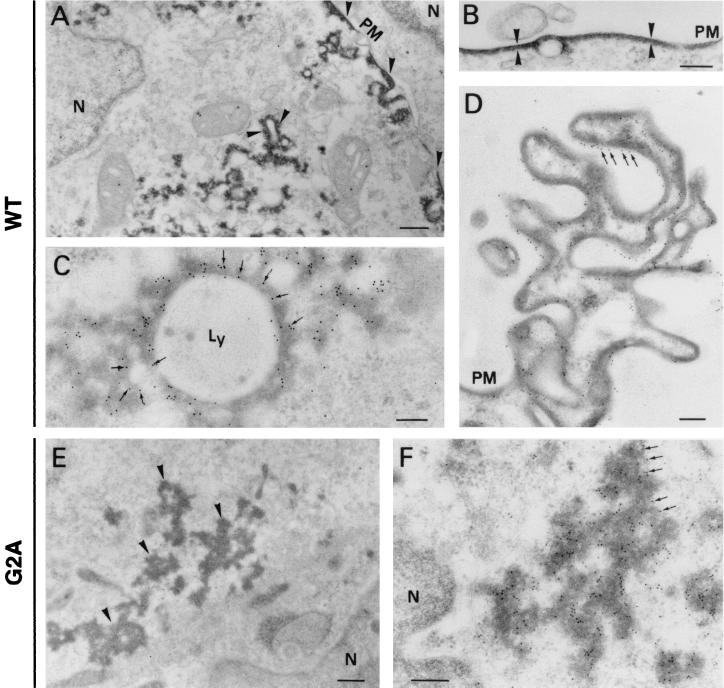
African swine fever virus (ASFV) polyprotein pp220, encoded by the CP2475L gene, is an N-myristoylated precursor polypeptide that, after proteolytic processing, gives rise to the major structural proteins p150, p37, p34, and p14. These proteins localize at the core shell, a matrix-like virus domain placed between the DNA-containing nucleoid and the inner envelope. In this study, we have examined the role of polyprotein pp220 in virus morphogenesis by means of an ASFV recombinant, v220i, containing an inducible copy of the CP2475L gene regulated by the Escherichia coli repressor-operator system. Under conditions that repress pp220 expression, the virus yield of v220i was about 2.6 log units lower than that of the parental virus or of the recombinant grown under permissive conditions. Electron microscopy revealed that pp220 repression leads to the assembly of icosahedral particles virtually devoid of the core structure. Analysis of recombinant v220i by immunoelectron microscopy, immunoblotting, and DNA hybridization showed that mutant particles essentially lack, besides the pp220-derived products, a number of major core proteins as well as the viral DNA. On the other hand, transient expression of the CP2475L gene in COS cells showed that polyprotein pp220 assembles into electron-dense membrane-bound coats, whereas a mutant nonmyristoylated version of pp220 does not associate with cellular membranes but forms large cytoplasmic aggregates. Together, these findings indicate that polyprotein pp220 is essential for the core assembly and suggest that its myristoyl moiety may function as a membrane-anchoring signal to bind the developing core shell to the inner viral envelope.
Figures
References
-
- Alves de Matos, A. P., and Z. G. Carvalho. 1993. African swine fever virus interaction with microtubules. Biol. Cell 78:229-234. - PubMed
-
- Andrés, G., A. Alejo, C. Simón-Mateo, and M. L. Salas. 2001. African swine fever virus protease: a new viral member of the SUMO-1-specific protease family. J. Biol. Chem. 276:780-787. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
