

CCR5 and CXCR4 usage by non-clade B human immunodeficiency virus type 1 primary isolates
- PMID: 11861874
- PMCID: PMC135972
- DOI: 10.1128/jvi.76.6.3059-3064.2002
CCR5 and CXCR4 usage by non-clade B human immunodeficiency virus type 1 primary isolates
Abstract
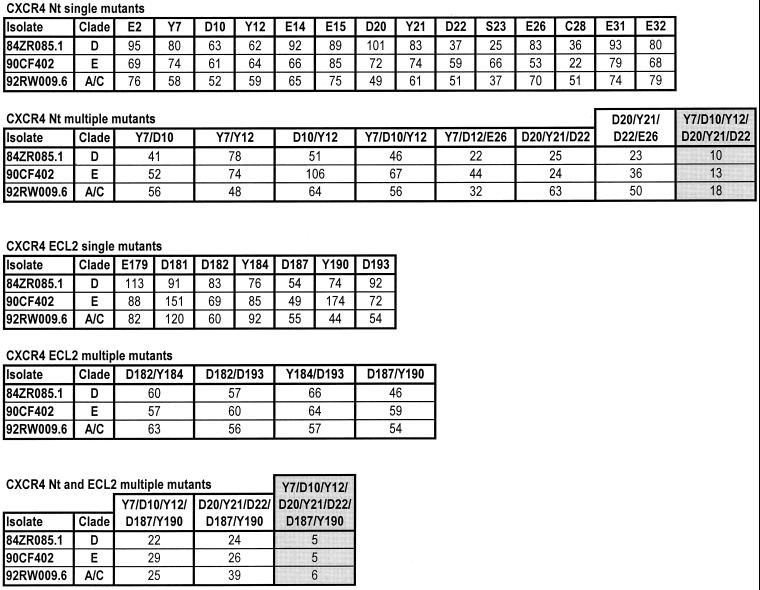
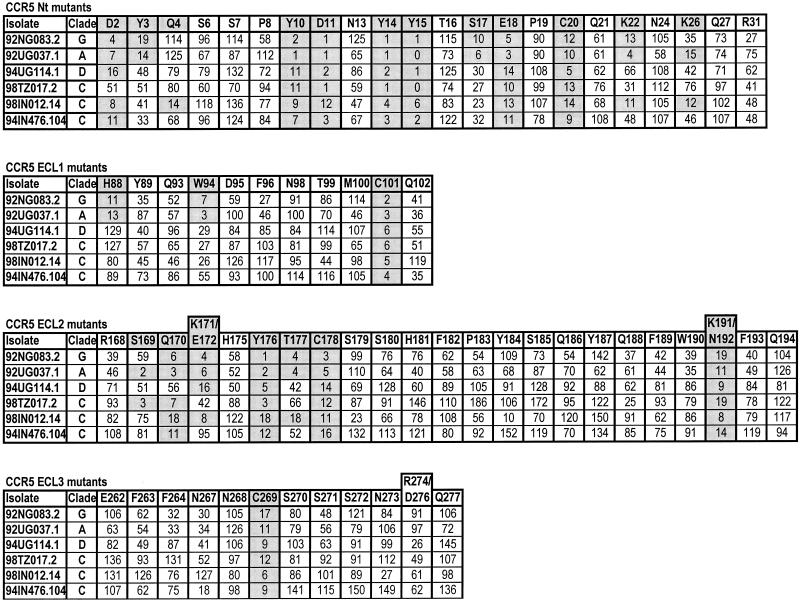
CCR5 and CXCR4 usage has been studied extensively with a variety of clade B human immunodeficiency virus type 1 (HIV-1) isolates. The determinants of CCR5 coreceptor function are remarkably consistent, with a region critical for fusion and entry located in the CCR5 amino-terminal domain (Nt). In particular, negatively charged amino acids and sulfated tyrosines in the Nt are essential for gp120 binding to CCR5. The same types of residues are important for CXCR4-mediated viral fusion and entry, but they are dispersed throughout the extracellular domains of CXCR4, and their usage is isolate dependent. Here, we report on the determinants of CCR5 and CXCR4 coreceptor function for a panel of non-clade B isolates that are responsible for the majority of new HIV-1 infections worldwide. Consistent with clade B isolates, CXCR4 usage remains isolate dependent and is determined by the overall content of negatively charged and tyrosine residues. Residues in the Nt of CCR5 that are important for fusion and entry of clade B isolates are also important for the entry of all non-clade B HIV-1 isolates that we tested. Surprisingly, we found that in contrast to clade B isolates, a cluster of residues in the second extracellular loop of CCR5 significantly affects fusion and entry of all non-clade B isolates tested. This points to a different mechanism of CCR5 usage by these viruses and may have important implications for the development of HIV-1 inhibitors that target CCR5 coreceptor function.
Figures

Similar articles
-
Tissue-specific sequence alterations in the human immunodeficiency virus type 1 envelope favoring CCR5 usage contribute to persistence of dual-tropic virus in the brain.J Virol. 2009 Jun;83(11):5430-41. doi: 10.1128/JVI.02648-08. Epub 2009 Mar 25. J Virol. 2009. PMID: 19321618 Free PMC article.
-
Role of the HIV type 1 glycoprotein 120 V3 loop in determining coreceptor usage.AIDS Res Hum Retroviruses. 1999 May 20;15(8):731-43. doi: 10.1089/088922299310827. AIDS Res Hum Retroviruses. 1999. PMID: 10357469
-
Sensitivity of human immunodeficiency virus type 1 to fusion inhibitors targeted to the gp41 first heptad repeat involves distinct regions of gp41 and is consistently modulated by gp120 interactions with the coreceptor.J Virol. 2001 Sep;75(18):8605-14. doi: 10.1128/jvi.75.18.8605-8614.2001. J Virol. 2001. PMID: 11507206 Free PMC article.
-
Effect of HIV-1 subtype and tropism on treatment with chemokine coreceptor entry inhibitors; overview of viral entry inhibition.Crit Rev Microbiol. 2015;41(4):473-87. doi: 10.3109/1040841X.2013.867829. Epub 2014 Mar 17. Crit Rev Microbiol. 2015. PMID: 24635642 Review.
-
Genotypic coreceptor analysis.Eur J Med Res. 2007 Oct 15;12(9):453-62. Eur J Med Res. 2007. PMID: 17933727 Review.
Cited by
-
Conserved determinants of enhanced CCR5 binding in the human immunodeficiency virus subtype D envelope third variable loop.AIDS Res Hum Retroviruses. 2010 Apr;26(4):481-8. doi: 10.1089/aid.2009.0086. AIDS Res Hum Retroviruses. 2010. PMID: 20377425 Free PMC article.
-
Potential Treatment of COVID-19 with Traditional Chinese Medicine: What Herbs Can Help Win the Battle with SARS-CoV-2?Engineering (Beijing). 2022 Dec;19:139-152. doi: 10.1016/j.eng.2021.08.020. Epub 2021 Oct 28. Engineering (Beijing). 2022. PMID: 34729244 Free PMC article. Review.
-
Direct observation to chemokine receptor 5 on T-lymphocyte cell surface using fluorescent metal nanoprobes.Biochem Biophys Res Commun. 2010 Sep 10;400(1):111-6. doi: 10.1016/j.bbrc.2010.08.020. Epub 2010 Aug 10. Biochem Biophys Res Commun. 2010. PMID: 20705055 Free PMC article.
-
The crown and stem of the V3 loop play distinct roles in human immunodeficiency virus type 1 envelope glycoprotein interactions with the CCR5 coreceptor.J Virol. 2002 Sep;76(17):8953-7. doi: 10.1128/jvi.76.17.8953-8957.2002. J Virol. 2002. PMID: 12163614 Free PMC article.
-
Allosteric model of maraviroc binding to CC chemokine receptor 5 (CCR5).J Biol Chem. 2011 Sep 23;286(38):33409-21. doi: 10.1074/jbc.M111.279596. Epub 2011 Jul 20. J Biol Chem. 2011. PMID: 21775441 Free PMC article.
References
-
- Bennetts, B. H., S. M. Teutsch, M. M. Buhler, R. N. Heard, and G. J. Stewart. 1997. The CCR5 deletion mutation fails to protect against multiple sclerosis. Hum. Immunol. 58:52-59. - PubMed
-
- Berger, E. 1997. HIV entry and tropism: the chemokine receptor expression. AIDS 11(Suppl. A):S3-S16. - PubMed
-
- Berger, E. A., P. M. Murphy, and J. M. Farber. 1999. Chemokine receptors as HIV-1 coreceptors: roles in viral entry, tropism, and disease. Annu. Rev. Immunol. 17:657-700. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
