

Phosphorylation of mammalian translation initiation factor 5 (eIF5) in vitro and in vivo
- PMID: 11861906
- PMCID: PMC101238
- DOI: 10.1093/nar/30.5.1154
Phosphorylation of mammalian translation initiation factor 5 (eIF5) in vitro and in vivo
Abstract
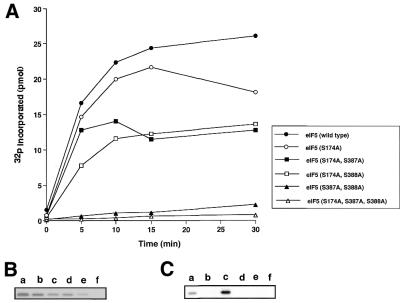
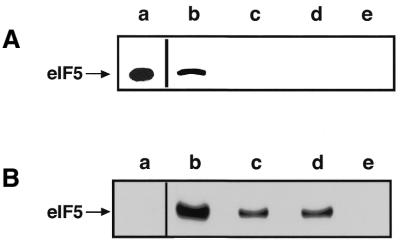
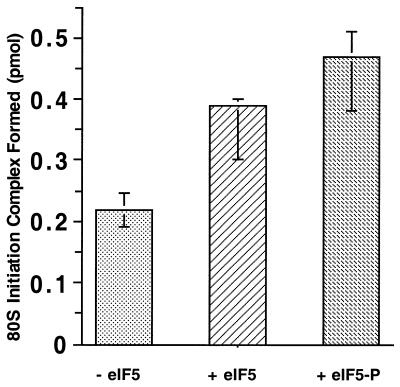
Eukaryotic translation initiation factor 5 (eIF5) interacts with the 40S initiation complex (40S*eIF3*AUG*Met-tRNA(f)*eIF2*GTP) and, acting as a GTPase activating protein, promotes the hydrolysis of bound GTP. We isolated a protein kinase from rabbit reticulocyte lysates on the basis of its ability to phosphorylate purified bacterially expressed recombinant rat eIF5. Physical, biochemical and antigenic properties of this kinase identify it as casein kinase II (CK II). Mass spectrometric analysis of maximally in vitro phosphorylated eIF5 localized the major phosphorylation sites at Ser-387 and Ser-388 near the C-terminus of eIF5. These serine residues are embedded within a cluster of acidic amino acid residues and account for nearly 90% of the total in vitro eIF5 phosphorylation. A minor phosphorylation site at Ser-174 was also observed. Alanine substitution mutagenesis at Ser-387 and Ser-388 of eIF5 abolishes phosphorylation by the purified kinase as well as by crude reticulocyte lysates. The same mutations also abolish phosphorylation of eIF5 when transfected into mammalian cells suggesting that CK II phosphorylates eIF5 at these two serine residues in vivo as well.
Figures

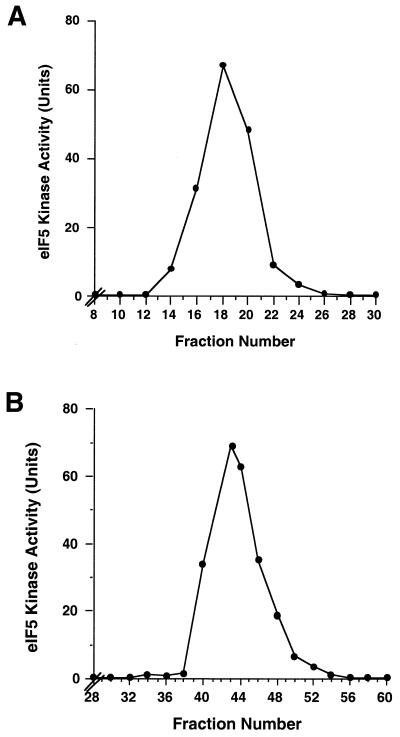
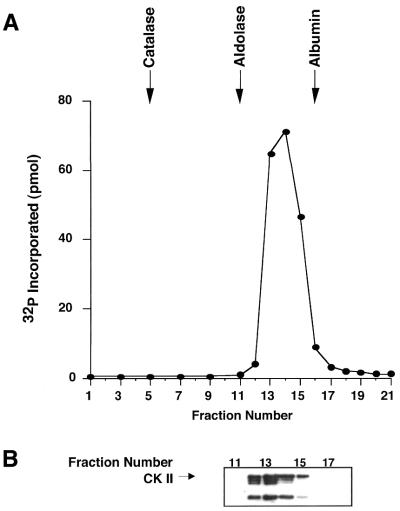
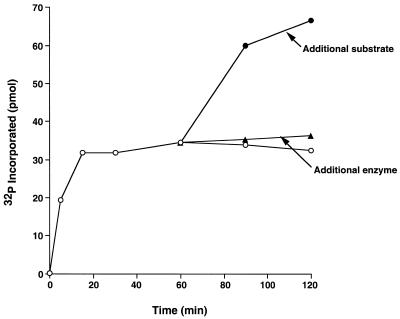
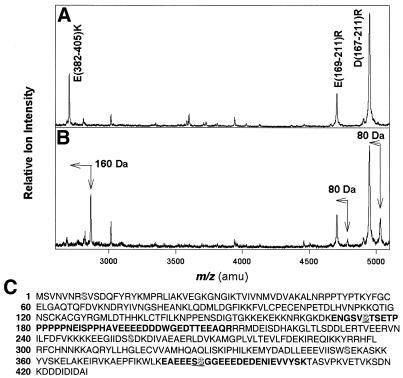
Similar articles
-
Casein kinase II phosphorylates translation initiation factor 5 (eIF5) in Saccharomyces cerevisiae.Yeast. 2003 Jan 30;20(2):97-108. doi: 10.1002/yea.937. Yeast. 2003. PMID: 12518314
-
Conserved bipartite motifs in yeast eIF5 and eIF2Bepsilon, GTPase-activating and GDP-GTP exchange factors in translation initiation, mediate binding to their common substrate eIF2.EMBO J. 1999 Mar 15;18(6):1673-88. doi: 10.1093/emboj/18.6.1673. EMBO J. 1999. PMID: 10075937 Free PMC article.
-
Mutational analysis of mammalian translation initiation factor 5 (eIF5): role of interaction between the beta subunit of eIF2 and eIF5 in eIF5 function in vitro and in vivo.Mol Cell Biol. 2000 Jun;20(11):3942-50. doi: 10.1128/MCB.20.11.3942-3950.2000. Mol Cell Biol. 2000. PMID: 10805737 Free PMC article.
-
Functional significance and mechanism of eIF5-promoted GTP hydrolysis in eukaryotic translation initiation.Prog Nucleic Acid Res Mol Biol. 2001;70:207-31. doi: 10.1016/s0079-6603(01)70018-9. Prog Nucleic Acid Res Mol Biol. 2001. PMID: 11642363 Review.
-
A multifactor complex of eukaryotic initiation factors, eIF1, eIF2, eIF3, eIF5, and initiator tRNA(Met) is an important translation initiation intermediate in vivo.Genes Dev. 2000 Oct 1;14(19):2534-46. doi: 10.1101/gad.831800. Genes Dev. 2000. PMID: 11018020 Free PMC article.
Cited by
-
Sliding of a 43S ribosomal complex from the recognized AUG codon triggered by a delay in eIF2-bound GTP hydrolysis.Nucleic Acids Res. 2016 Feb 29;44(4):1882-93. doi: 10.1093/nar/gkv1514. Epub 2015 Dec 29. Nucleic Acids Res. 2016. PMID: 26717981 Free PMC article.
-
Early cytokinin response proteins and phosphoproteins of Arabidopsis thaliana identified by proteome and phosphoproteome profiling.J Exp Bot. 2011 Jan;62(3):921-37. doi: 10.1093/jxb/erq322. Epub 2010 Oct 25. J Exp Bot. 2011. PMID: 20974740 Free PMC article.
-
Chemical Genetic Validation of CSNK2 Substrates Using an Inhibitor-Resistant Mutant in Combination with Triple SILAC Quantitative Phosphoproteomics.Front Mol Biosci. 2022 Jun 9;9:909711. doi: 10.3389/fmolb.2022.909711. eCollection 2022. Front Mol Biosci. 2022. PMID: 35755813 Free PMC article.
-
Phosphorylation of plant translation initiation factors by CK2 enhances the in vitro interaction of multifactor complex components.J Biol Chem. 2009 Jul 31;284(31):20615-28. doi: 10.1074/jbc.M109.007658. Epub 2009 Jun 9. J Biol Chem. 2009. PMID: 19509420 Free PMC article.
-
The N-terminal domain of the human eIF2beta subunit and the CK2 phosphorylation sites are required for its function.Biochem J. 2006 Feb 15;394(Pt 1):227-36. doi: 10.1042/BJ20050605. Biochem J. 2006. PMID: 16225457 Free PMC article.
References
-
- Raychaudhuri P., Chaudhuri,A. and Maitra,U. (1985) Eukaryotic initiation factor 5 from calf liver is a single polypeptide chain protein of Mr = 62,000. J. Biol. Chem., 260, 2132–2139. - PubMed
-
- Chevesich J., Chaudhuri,J. and Maitra,U. (1993) Characterization of mammalian translation initiation factor 5 (eIF-5). Demonstration that eIF-5 is a phosphoprotein and is present in cells as a single molecular form of apparent Mr 58,000. J. Biol. Chem., 268, 20659–20667. - PubMed
-
- Chakravarti D., Maiti,T. and Maitra,U. (1993) Isolation and immunochemical characterization of eukaryotic translation initiation factor 5 from Saccharomyces cerevisiae. J. Biol. Chem., 268, 5754–5762. - PubMed
-
- Chakravarti D. and Maitra,U. (1993) Eukaryotic translation initiation factor 5 from Saccharomyces cerevisiae. Cloning, characterization and expression of the gene encoding the 45,346-Da protein. J. Biol. Chem., 268, 10524–10533. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
