

RNA destabilization by the granulocyte colony-stimulating factor stem-loop destabilizing element involves a single stem-loop that promotes deadenylation
- PMID: 11865046
- PMCID: PMC135610
- DOI: 10.1128/MCB.22.6.1664-1673.2002
RNA destabilization by the granulocyte colony-stimulating factor stem-loop destabilizing element involves a single stem-loop that promotes deadenylation
Abstract
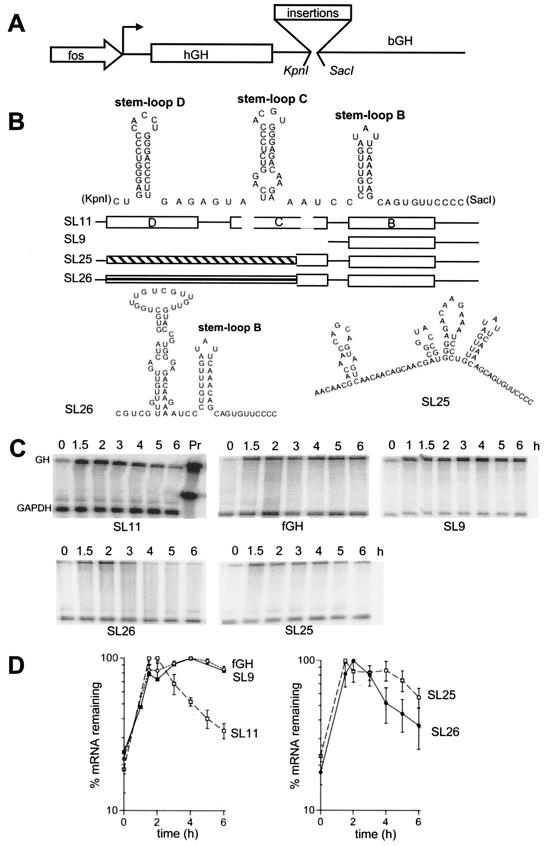
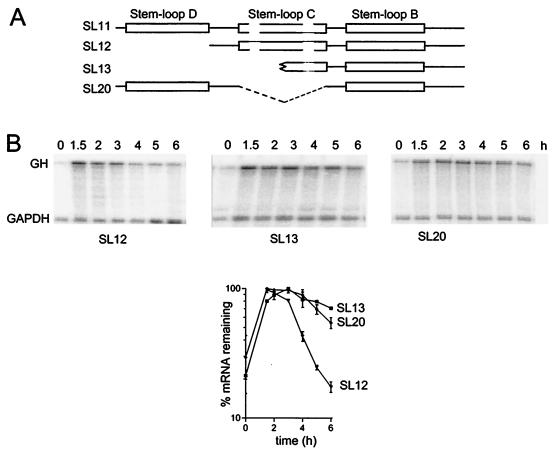
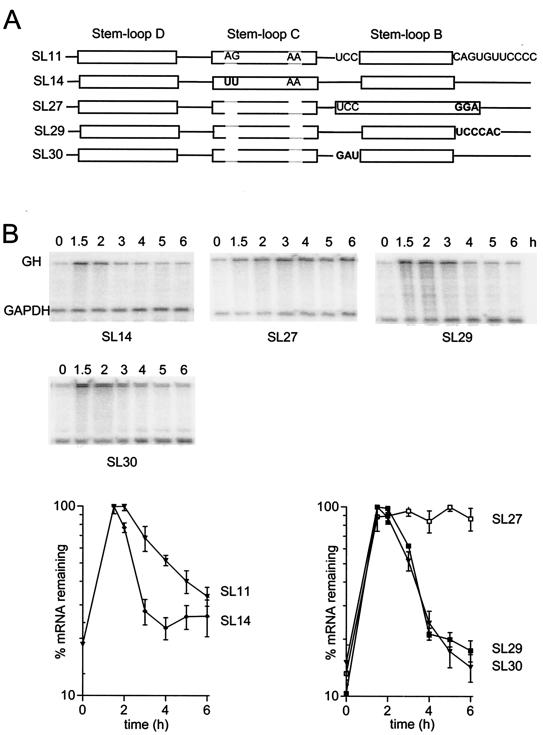
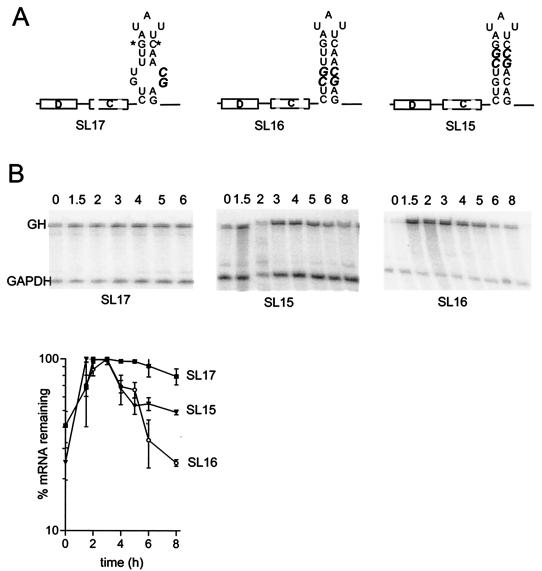
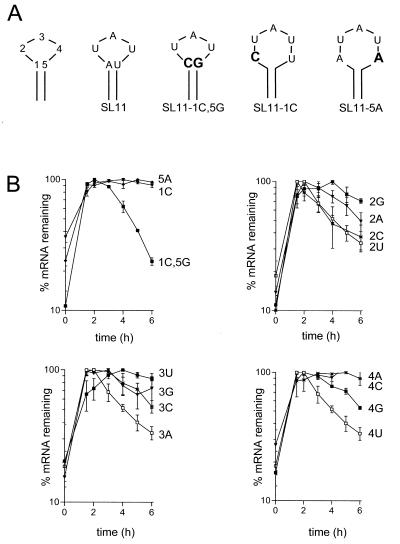
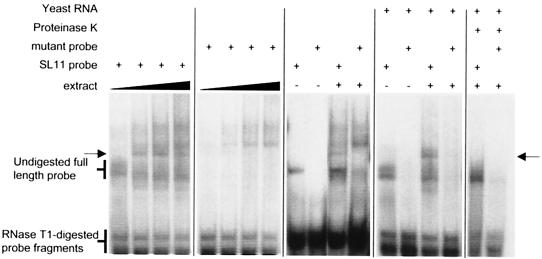
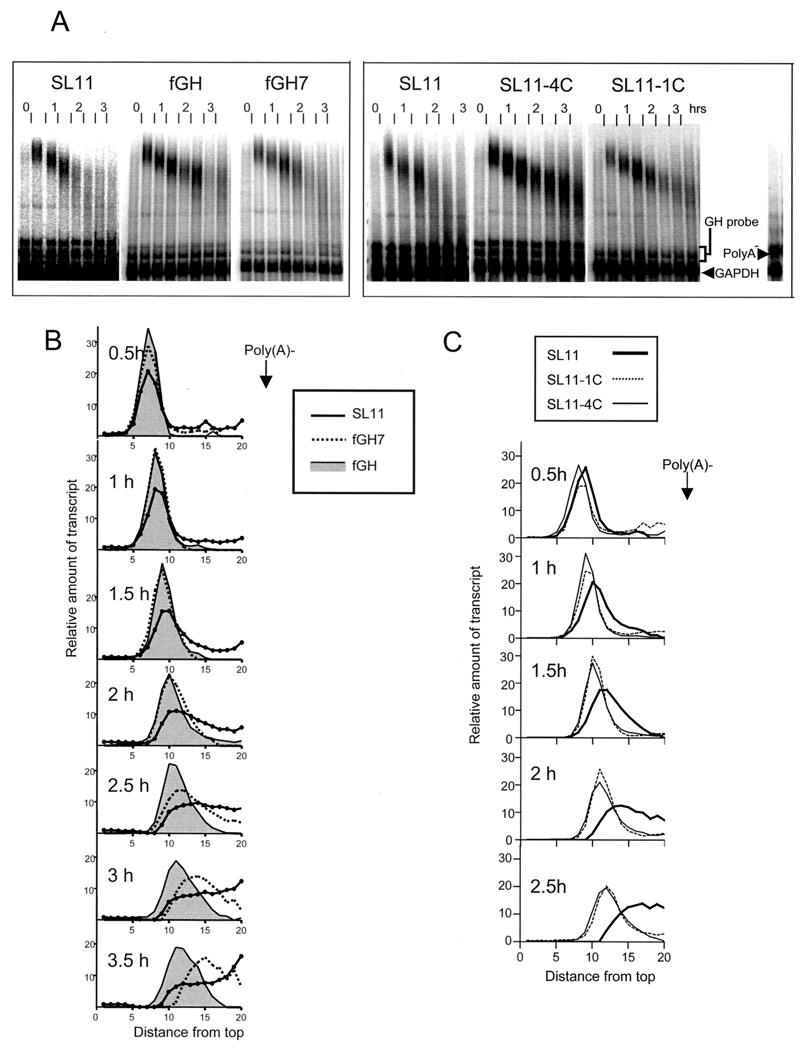
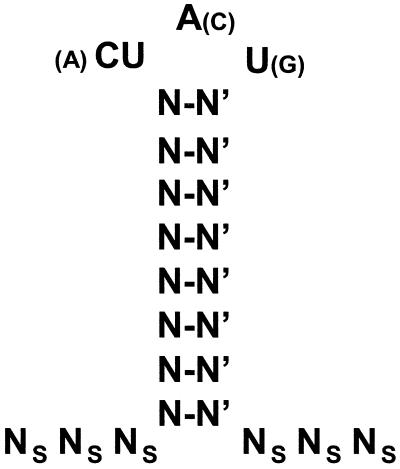
Granulocyte colony-stimulating factor (G-CSF) mRNA contains two distinct types of cis-acting mRNA destabilizing elements in the 3'-untranslated region. In addition to several copies of the AU-rich element the G-CSF mRNA also contains a destabilizing region that includes several predicted stem-loop structures. We report here that the destabilizing activity resides in a single stem-loop structure within this region. A consensus sequence for the active structure has been derived by site-directed mutagenesis, revealing that a three-base loop of sequence YAU and unpaired bases either side of the stem contribute to the activity. The helical nature of the stem is essential and the stem must be less than 11 bp in length, but the destabilizing activity is relatively insensitive to the sequence within the helix. The stem-loop increases the rate of mRNA deadenylation, most likely by enhancing the processivity of the deadenylation reaction. A protein that binds the stem-loop, but not an inactive mutant form, has been detected in cytoplasmic lysates.
Figures

References
-
- Binder, R., J. A. Horowitz, J. P. Basilion, D. M. Koeller, R. D. Klausner, and J. B. Harford. 1994. Evidence that the pathway of transferrin receptor mRNA degradation involves an endonucleolytic cleavage within the 3′ UTR and does not involve poly(A) tail shortening. EMBO J. 13:1969-1980. - PMC - PubMed
-
- Binder, R., S.-P. L. Hwang, R. Ratnasabapathy, and D. L. Williams. 1989. Degradation of apolipoprotein II mRNA occurs via endonucleolytic cleavage at 5′-AAU-3′/5/-UAA-3′ elements in single-stranded loop domains of the 3′-noncoding region. J. Biol. Chem. 264:16910-16918. - PubMed
-
- Brown, B. D., I. D. Zipkin, and R. M. Harland. 1993. Sequence-specific endonucleolytic cleavage and protection of mRNA in Xenopus and Drosophila. Genes Dev. 7:1620-1631. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases