

Caspase cleavage of initiation factor 4E-binding protein 1 yields a dominant inhibitor of cap-dependent translation and reveals a novel regulatory motif
- PMID: 11865047
- PMCID: PMC135612
- DOI: 10.1128/MCB.22.6.1674-1683.2002
Caspase cleavage of initiation factor 4E-binding protein 1 yields a dominant inhibitor of cap-dependent translation and reveals a novel regulatory motif
Abstract
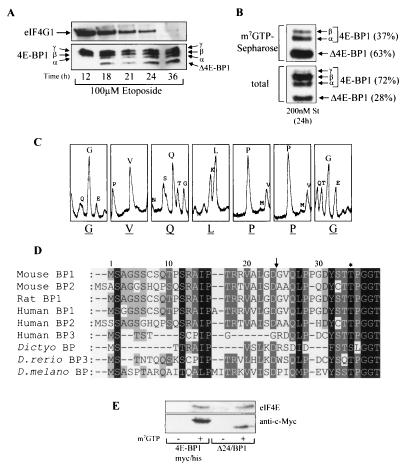
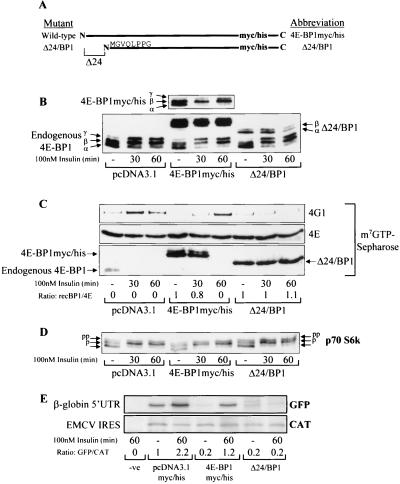
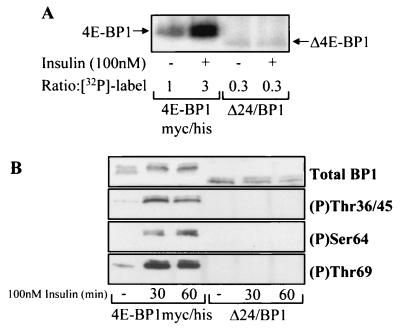
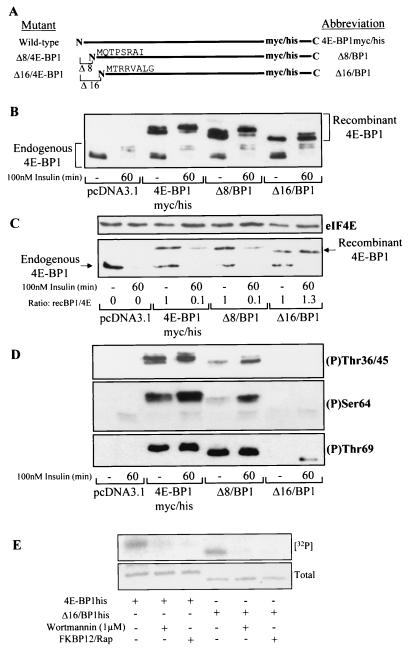
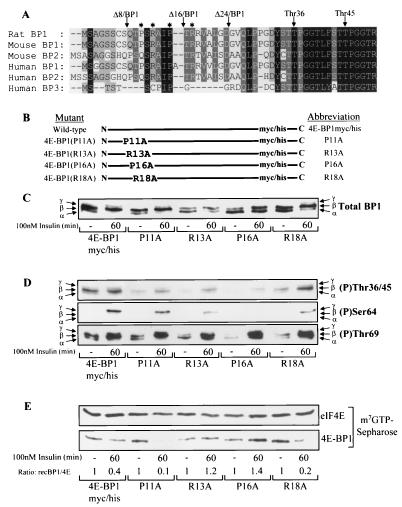
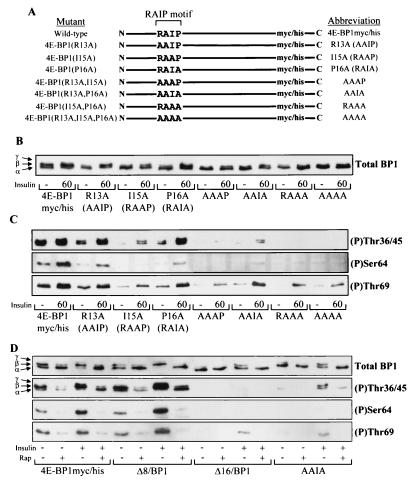
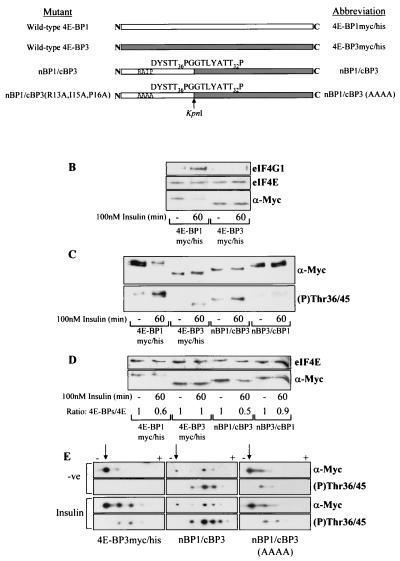
Eukaryotic initiation factor 4E (eIF4E) binding proteins (4E-BPs) regulate the assembly of initiation complexes required for cap-dependent mRNA translation. 4E-BP1 undergoes insulin-stimulated phosphorylation, resulting in its release from eIF4E, allowing initiation complex assembly. 4E-BP1 undergoes caspase-dependent cleavage in cells undergoing apoptosis. Here we show that cleavage occurs after Asp24, giving rise to the N-terminally truncated polypeptide Delta4E-BP1, which possesses the eIF4E-binding site and all the known phosphorylation sites. Delta4E-BP1 binds to eIF4E and fails to become sufficiently phosphorylated upon insulin stimulation to bring about its release from eIF4E. Therefore, Delta4E-BP1 acts as a potent inhibitor of cap-dependent translation. Using a mutagenesis approach, we identify a novel regulatory motif of four amino acids (RAIP) which lies within the first 24 residues of 4E-BP1 and which is necessary for efficient phosphorylation of 4E-BP1. This motif is conserved among sequences of 4E-BP1 and 4E-BP2 but is absent from 4E-BP3. Insulin increased the phosphorylation of 4E-BP3 but not sufficiently to cause its release from eIF4E. However, a chimeric protein that was generated by replacing the N terminus of 4E-BP3 with the N-terminal sequence of 4E-BP1 (containing this RAIP motif) underwent a higher degree of phosphorylation and was released from eIF4E. This suggests that the N-terminal sequence of 4E-BP1 is required for optimal regulation of 4E-BPs by insulin.
Figures
References
-
- Brunn, G. J., C. C. Hudson, A. Sekulic, J. M. Williams, H. Hosoi, P. J. Houghton, J. C. Lawrence, and R. T. Abraham. 1997. Phosphorylation of the translational repressor PHAS-I by the mammalian target of rapamycin. Science 277:99-101. - PubMed
-
- Bushell, M., L. McKendrick, R. U. Jaenicke, M. J. Clemens, and S. J. Morley. 1999. Caspase-3 is necessary and sufficient for cleavage of protein synthesis eukaryotic initiation factor 4G during apoptosis. FEBS Lett. 451:332-336. - PubMed
-
- Bushell, M., D. Poncet, W. E. Marissen, H. Flotow, R. E. Lloyd, M. J. Clemens, and S. J. Morley. 2000. Cleavage of polypeptide chain initiation factor eIF4GI during apoptosis in lymphoma cells: characterisation of an internal fragment by caspase-3-mediated cleavage. Cell Death Differ. 7:628-636. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous