

The morphogenic linker peptide of HBV capsid protein forms a mobile array on the interior surface
- PMID: 11867516
- PMCID: PMC125892
- DOI: 10.1093/emboj/21.5.876
The morphogenic linker peptide of HBV capsid protein forms a mobile array on the interior surface
Abstract
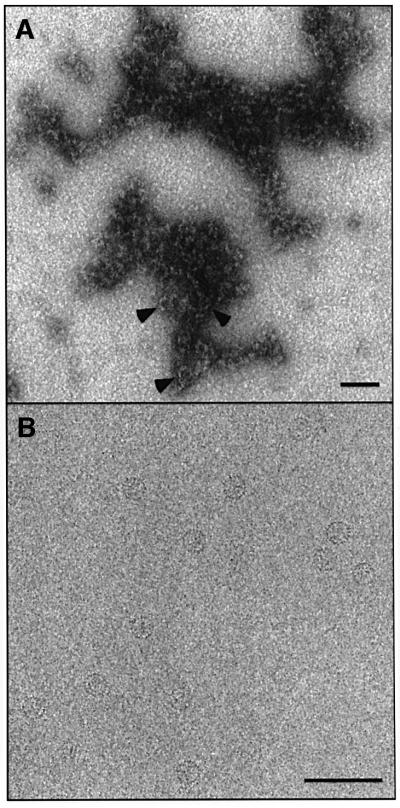
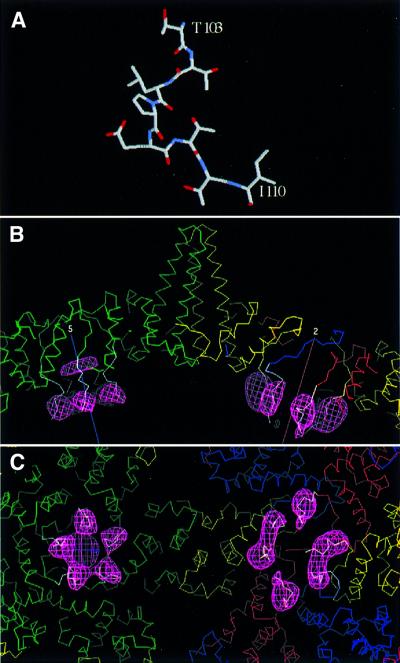
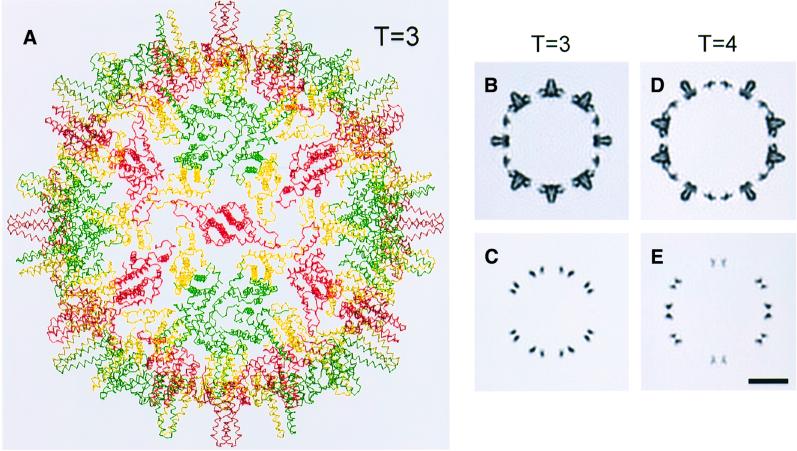
Many capsid proteins have peptides that influence their assembly. In hepatitis B virus capsid protein, the peptide STLPETTVV, linking the shell-forming 'core' domain and the nucleic acid-binding 'protamine' domain, has such a role. We have studied its morphogenic properties by permuting its sequence, substituting it with an extraneous peptide, deleting it to directly fuse the core and protamine domains and assembling core domain dimers with added linker peptides. The peptide was found to be necessary for the assembly of protamine domain-containing capsids, although its size-determining effect tolerates some modifications. Although largely invisible in a capsid crystal structure, we could visualize linker peptides by cryo-EM difference imaging: they emerge on the inner surface and extend from the capsid protein dimer interface towards the adjacent symmetry axis. A closely sequence-similar peptide in cellobiose dehydrogenase, which has an extended conformation, offers a plausible prototype. We propose that linker peptides are attached to the capsid inner surface as hinged struts, forming a mobile array, an arrangement with implications for morphogenesis and the management of encapsidated nucleic acid.
Figures
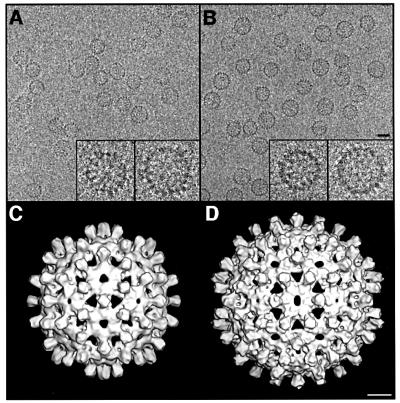
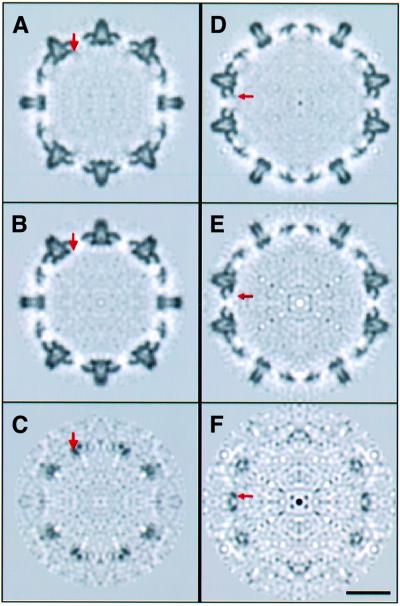
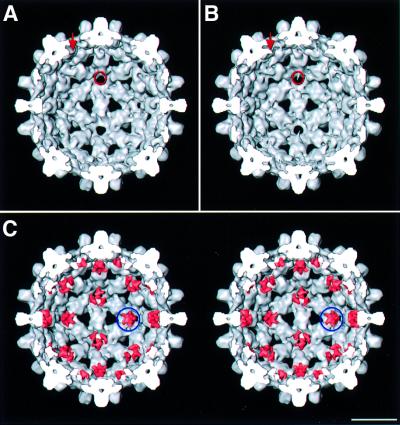
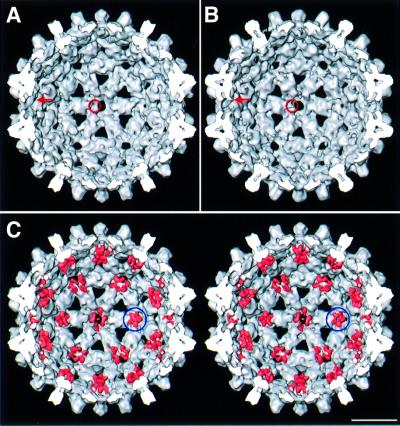
References
-
- Baker T.S. and Cheng,R.H. (1996) A model-based approach for determining orientations of biological macromolecules imaged by cryoelectron microscopy. J. Struct. Biol., 116, 120–130. - PubMed
-
- Beames B. and Lanford,R.E. (1993) Carboxy-terminal truncations of the HBV core protein affect capsid formation and the apparent size of encapsidated HBV RNA. Virology, 194, 597–607. - PubMed
-
- Belnap D.M., Kumar,A., Folk,J.T., Smith,T.J. and Baker,T.S. (1999) Low-resolution density maps from atomic models: how stepping ‘back’ can be a step ‘forward’. J. Struct. Biol., 125, 166–175. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
