

The Tim9p-Tim10p complex binds to the transmembrane domains of the ADP/ATP carrier
- PMID: 11867522
- PMCID: PMC125908
- DOI: 10.1093/emboj/21.5.942
The Tim9p-Tim10p complex binds to the transmembrane domains of the ADP/ATP carrier
Abstract
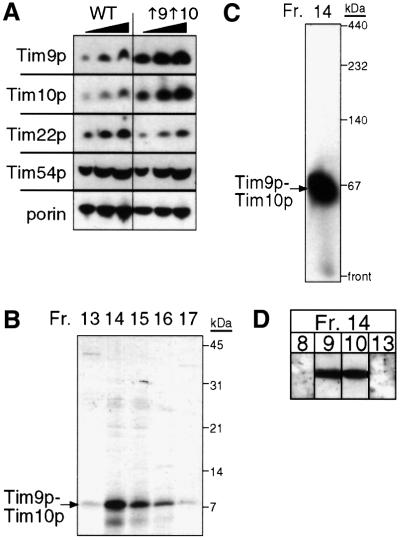
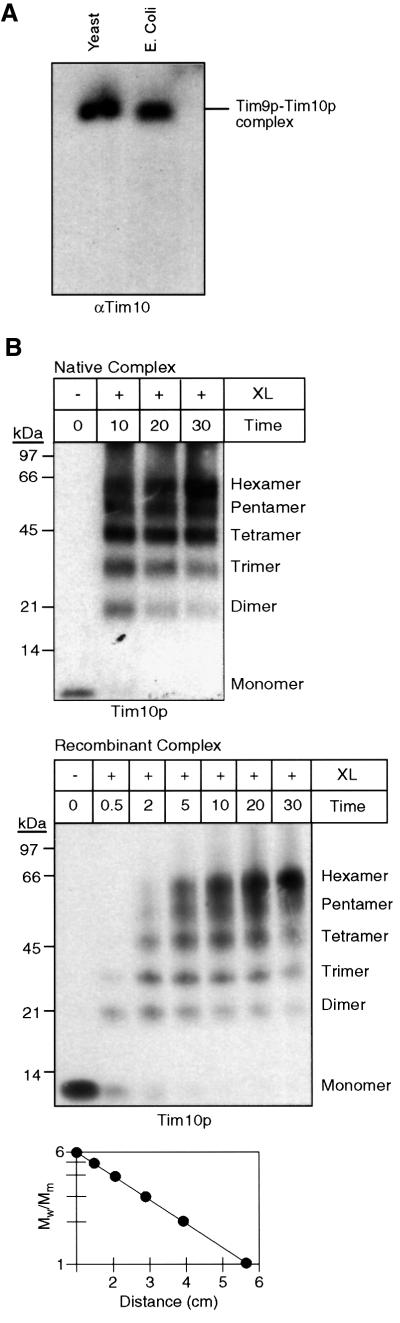
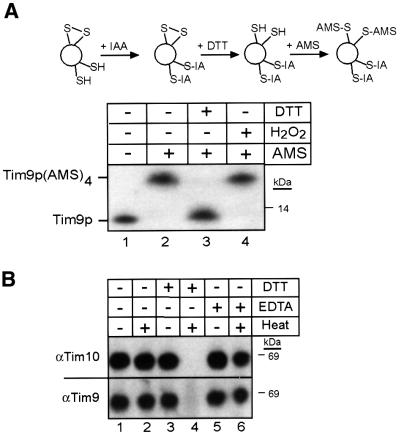
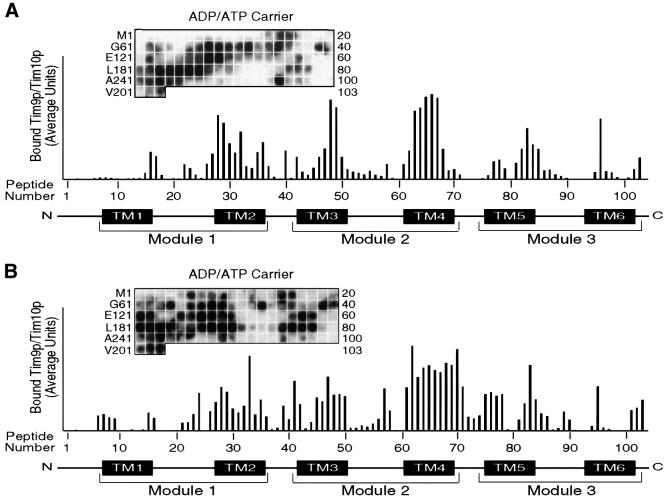
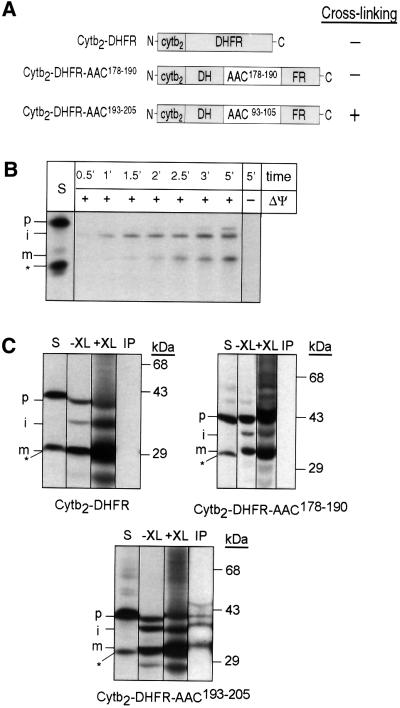
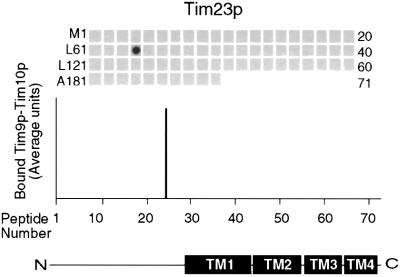
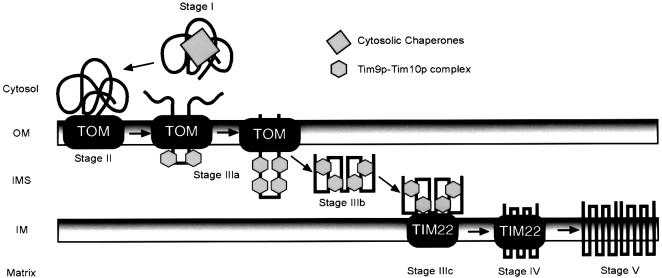
The soluble Tim9p-Tim10p (Tim, translocase of inner membrane) complex of the mitochondrial intermembrane space mediates the import of the carrier proteins and is a component of the TIM22 import system. The mechanism by which the Tim9p-Tim10p complex assembles and binds the carriers is not well understood, but previous studies have proposed that the conserved cysteine residues in the 'twin CX3C' motif coordinate zinc and potentially generate a zinc-finger-like structure that binds to the matrix loops of the carrier proteins. Here we have purified the native and recombinant Tim9p-Tim10p complex, and show that both complexes resemble each other and consist of three Tim9p and three Tim10p. Results from inductively coupled plasma--mass spectrometry studies failed to detect zinc in the Tim9p-Tim10p complex. Instead, the cysteine residues seemingly formed disulfide linkages. The Tim9p-Tim10p complex bound specifically to the transmembrane domains of the ADP/ATP carrier, but had no affinity for Tim23p, an inner membrane protein that is inserted via the TIM22 complex. The chaperone-like Tim9p-Tim10p complex thus may prevent aggregation of the unfolded carrier proteins in the aqueous intermembrane space.
Figures
References
-
- Azem A., Weiss,C. and Goloubinoff,P. (1998) Structural analysis of GroE chaperonin complexes using chemical cross-linking. Methods Enzymol., 290, 253–268. - PubMed
-
- Bauer M.F., Hofmann,S., Neupert,W. and Brunner,M. (2000) Protein translocation into mitochondria: the role of TIM complexes. Trends Cell Biol., 10, 25–31. - PubMed
-
- Brix J., Rudiger,S., Bukau,B., Schneider-Mergener,J. and Pfanner,N. (1999) Distribution of binding sequences for the mitochondrial import receptors Tom20, Tom22 and Tom70 in a presequence-carrying preprotein and a non-cleavable preprotein. J. Biol. Chem., 274, 16522–16530. - PubMed
-
- Brix J., Ziegler,G.A., Dietmeier,K., Schneider-Mergener,J., Schulz,G.E. and Pfanner,N. (2000) The mitochondrial import receptor Tom70: identification of a 25 kDa core domain with a specific binding site for preproteins. J. Mol. Biol., 303, 479–488. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
