

Visualization of the ER-to-cytosol dislocation reaction of a type I membrane protein
- PMID: 11867532
- PMCID: PMC125905
- DOI: 10.1093/emboj/21.5.1041
Visualization of the ER-to-cytosol dislocation reaction of a type I membrane protein
Abstract
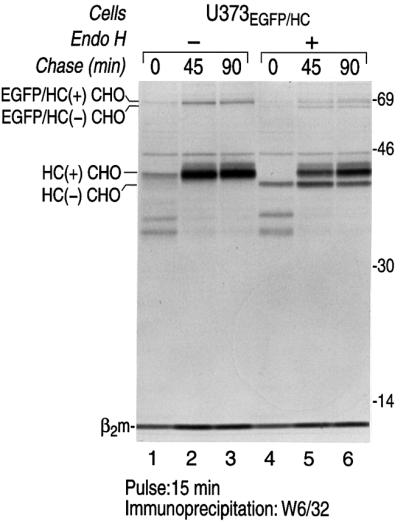
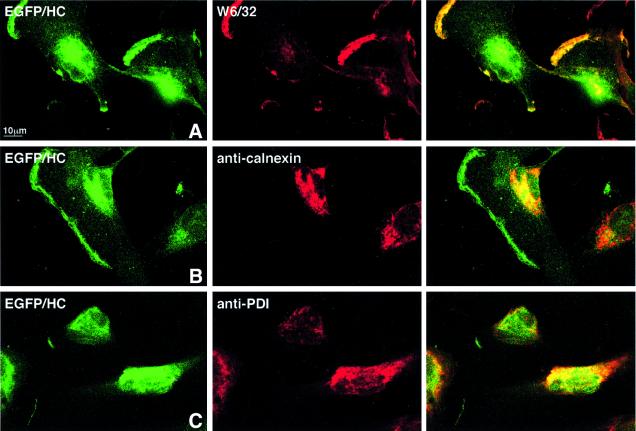
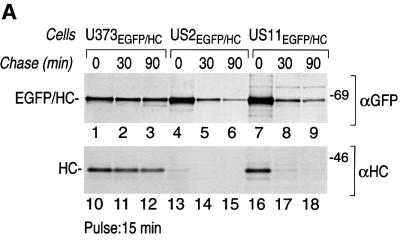
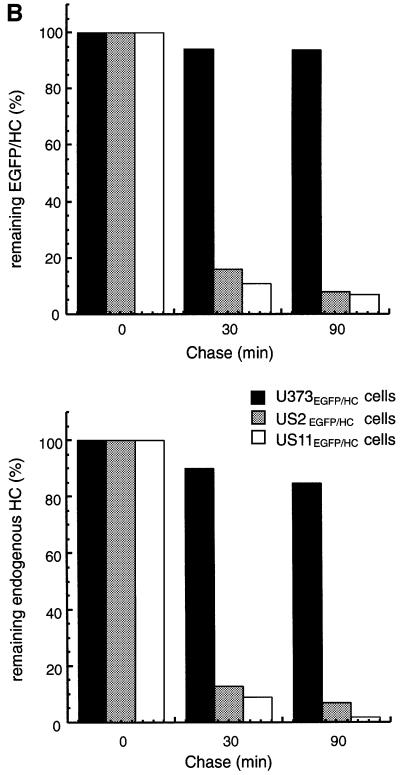
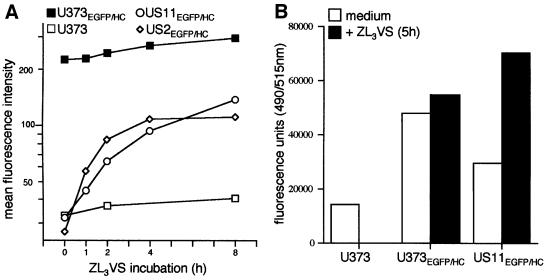
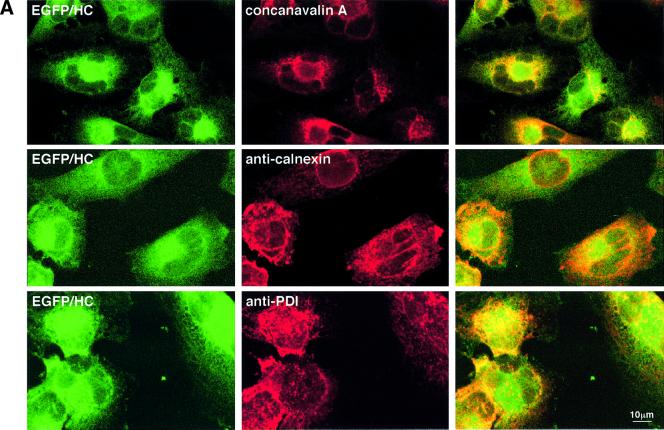
The human cytomegalovirus gene products US2 and US11 induce proteasomal degradation of MHC class I heavy chains. We have generated an enhanced green fluorescent protein-class I heavy chain (EGFP-HC) chimeric molecule to study its dislocation and degradation in US2- and US11-expressing cells. The EGFP-HC fusion is stable in control cells, but is degraded rapidly in US2- or US11-expressing cells. Proteasome inhibitors induce in a time-dependent manner the accumulation of EGFP-HC molecules in US2- and US11-expressing cells, as assessed biochemically and by cytofluorimetry of intact cells. Pulse-chase analysis and subcellular fractionation show that EGFP-HC proteins are dislocated from the endoplasmic reticulum and can be recovered as deglycosylated fluorescent intermediates in the cytosol. These results raise the possibility that dislocation of glycoproteins from the ER may not require their full unfolding.
Figures







Similar articles
-
TRAM1 participates in human cytomegalovirus US2- and US11-mediated dislocation of an endoplasmic reticulum membrane glycoprotein.J Biol Chem. 2009 Feb 27;284(9):5905-14. doi: 10.1074/jbc.M807568200. Epub 2009 Jan 2. J Biol Chem. 2009. PMID: 19121997 Free PMC article.
-
Membrane-specific, host-derived factors are required for US2- and US11-mediated degradation of major histocompatibility complex class I molecules.J Biol Chem. 2002 Feb 1;277(5):3258-67. doi: 10.1074/jbc.M109765200. Epub 2001 Nov 20. J Biol Chem. 2002. PMID: 11717308
-
The cytosolic tail of class I MHC heavy chain is required for its dislocation by the human cytomegalovirus US2 and US11 gene products.Proc Natl Acad Sci U S A. 1999 Jul 20;96(15):8516-21. doi: 10.1073/pnas.96.15.8516. Proc Natl Acad Sci U S A. 1999. PMID: 10411907 Free PMC article.
-
The HCMV gene products US2 and US11 target MHC class I molecules for degradation in the cytosol.Curr Top Microbiol Immunol. 2002;269:37-55. doi: 10.1007/978-3-642-59421-2_3. Curr Top Microbiol Immunol. 2002. PMID: 12224515 Review.
-
Endoplasmic reticulum-associated protein degradation.Semin Cell Dev Biol. 2000 Jun;11(3):159-64. doi: 10.1006/scdb.2000.0160. Semin Cell Dev Biol. 2000. PMID: 10906272 Review.
Cited by
-
A glycosylated type I membrane protein becomes cytosolic when peptide: N-glycanase is compromised.EMBO J. 2004 Feb 11;23(3):650-8. doi: 10.1038/sj.emboj.7600090. Epub 2004 Jan 29. EMBO J. 2004. PMID: 14749736 Free PMC article.
-
Viral and cellular MARCH ubiquitin ligases and cancer.Semin Cancer Biol. 2008 Dec;18(6):441-50. doi: 10.1016/j.semcancer.2008.09.002. Epub 2008 Oct 2. Semin Cancer Biol. 2008. PMID: 18948196 Free PMC article. Review.
-
Roles of p97-associated deubiquitinases in protein quality control at the endoplasmic reticulum.Curr Protein Pept Sci. 2012 Aug;13(5):436-46. doi: 10.2174/138920312802430608. Curr Protein Pept Sci. 2012. PMID: 22812527 Free PMC article. Review.
-
The delicate balance between secreted protein folding and endoplasmic reticulum-associated degradation in human physiology.Physiol Rev. 2012 Apr;92(2):537-76. doi: 10.1152/physrev.00027.2011. Physiol Rev. 2012. PMID: 22535891 Free PMC article. Review.
-
Doa10 is a membrane protein retrotranslocase in ER-associated protein degradation.Elife. 2020 Jun 26;9:e56945. doi: 10.7554/eLife.56945. Elife. 2020. PMID: 32588820 Free PMC article.
References
-
- Battistutta R., Negro,A. and Zanotti,G. (2000) Crystal structure and refolding properties of the mutant F99S/M153T/V163A of the green fluorescent protein. Proteins, 41, 429–437. - PubMed
-
- Bebok Z., Mazzochi,C., King,S.A., Hong,J.S. and Sorscher,E.J. (1998) The mechanism underlying cystic fibrosis transmembrane conductance regulator transport from the endoplasmic reticulum to the proteasome includes Sec61β and a cytosolic, deglycosylated intermediary. J. Biol. Chem., 273, 29873–29878. - PubMed
-
- Beckmann R., Spahn,C.M., Eswar,N., Helmers,J., Penczek,P.A., Sali,A., Frank,J. and Blobel,G. (2001) Architecture of the protein-conducting channel associated with the translating 80S ribosome. Cell, 107, 361–372. - PubMed
-
- Biederer T., Volkwein,C. and Sommer,T. (1997) Role of Cue1p in ubiquitination and degradation at the ER surface. Science, 278, 1806–1809. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
