

Drosophila Gp150 is required for early ommatidial development through modulation of Notch signaling
- PMID: 11867535
- PMCID: PMC125890
- DOI: 10.1093/emboj/21.5.1074
Drosophila Gp150 is required for early ommatidial development through modulation of Notch signaling
Abstract
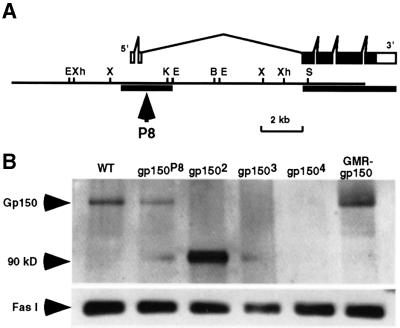
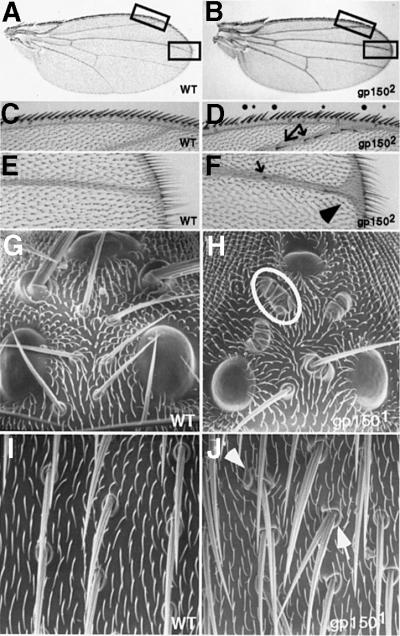
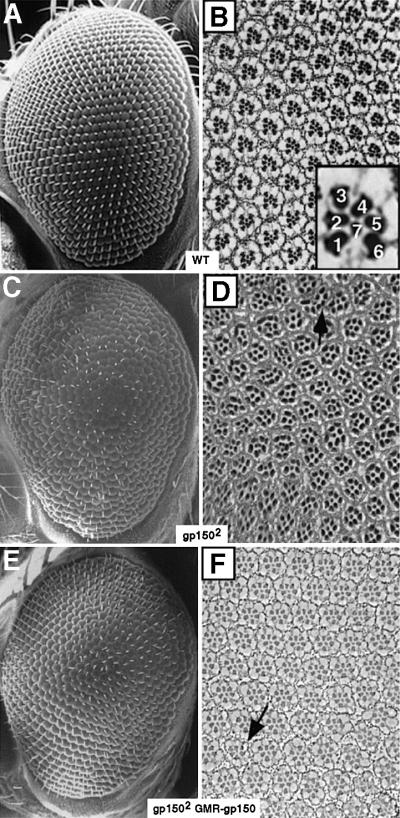
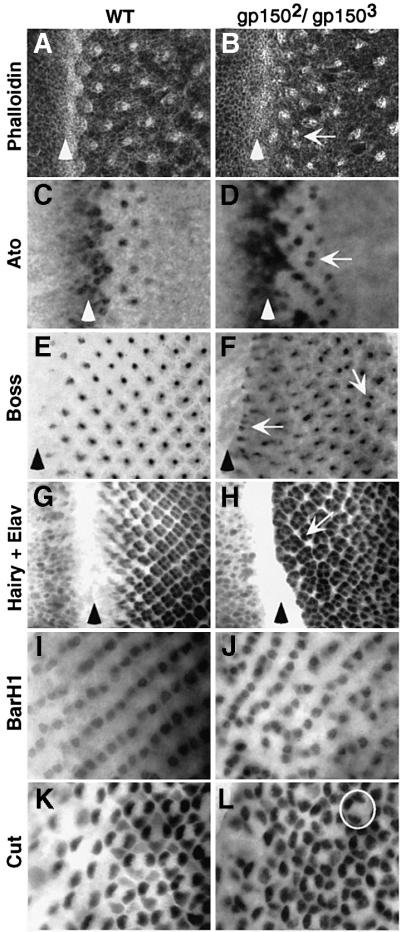
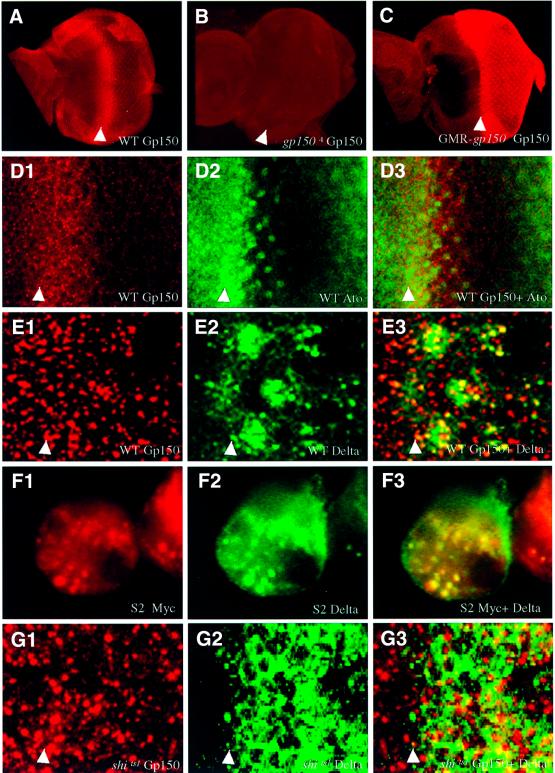
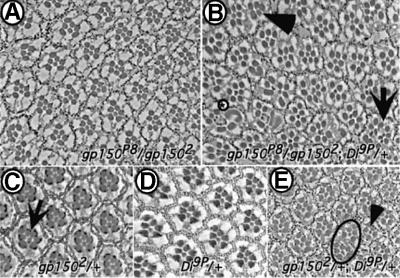
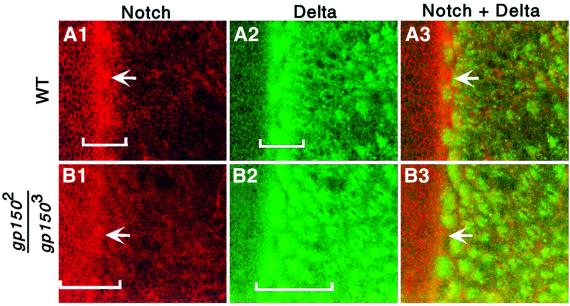
Cellular signaling activities must be tightly regulated for proper cell fate control and tissue morphogenesis. Here we report that the Drosophila leucine-rich repeat transmembrane glycoprotein Gp150 is required for viability, fertility and development of the eye, wing and sensory organs. In the eye, Gp150 plays a critical role in regulating early ommatidial formation. Gp150 is highly expressed in cells of the morphogenetic furrow (MF) region, where it accumulates exclusively in intracellular vesicles in an endocytosis-independent manner. Loss of gp150 function causes defects in the refinement of photoreceptor R8 cells and recruitment of other cells, which leads to the formation of aberrant ommatidia. Genetic analyses suggest that Gp150 functions to modulate Notch signaling. Consistent with this notion, Gp150 is co-localized with Delta in intracellular vesicles in cells within the MF region and loss of gp150 function causes accumulation of intracellular Delta protein. Therefore, Gp150 might function in intracellular vesicles to modulate Delta-Notch signaling for cell fate control and tissue morphogenesis.
Figures
Similar articles
-
Scabrous and Gp150 are endosomal proteins that regulate Notch activity.Development. 2003 Jul;130(13):2819-27. doi: 10.1242/dev.00495. Development. 2003. PMID: 12756167
-
The R8-photoreceptor equivalence group in Drosophila: fate choice precedes regulated Delta transcription and is independent of Notch gene dose.Mech Dev. 1998 Jun;74(1-2):3-14. doi: 10.1016/s0925-4773(98)00054-9. Mech Dev. 1998. PMID: 9651468
-
canoe encodes a novel protein containing a GLGF/DHR motif and functions with Notch and scabrous in common developmental pathways in Drosophila.Genes Dev. 1995 Mar 1;9(5):612-25. doi: 10.1101/gad.9.5.612. Genes Dev. 1995. PMID: 7698650
-
Delta-notch signaling and Drosophila cell fate choice.Dev Biol. 1994 Dec;166(2):415-30. doi: 10.1006/dbio.1994.1326. Dev Biol. 1994. PMID: 7813766 Review.
-
Signal transduction and the fate of the R7 photoreceptor in Drosophila.Trends Genet. 1991 Nov-Dec;7(11-12):372-7. doi: 10.1016/0168-9525(91)90258-r. Trends Genet. 1991. PMID: 1668189 Review.
Cited by
-
A conserved major facilitator superfamily member orchestrates a subset of O-glycosylation to aid macrophage tissue invasion.Elife. 2019 Mar 26;8:e41801. doi: 10.7554/eLife.41801. Elife. 2019. PMID: 30910009 Free PMC article.
-
The extracellular leucine-rich repeat superfamily; a comparative survey and analysis of evolutionary relationships and expression patterns.BMC Genomics. 2007 Sep 14;8:320. doi: 10.1186/1471-2164-8-320. BMC Genomics. 2007. PMID: 17868438 Free PMC article.
-
Competition between Delta and the Abruptex domain of Notch.BMC Dev Biol. 2008 Jan 21;8:4. doi: 10.1186/1471-213X-8-4. BMC Dev Biol. 2008. PMID: 18208612 Free PMC article.
References
-
- Artavanis-Tsakonas S., Rand,M.D. and Lake,R.J. (1999) Notch signaling: cell fate control and signal integration in development. Science, 284, 770–776. - PubMed
-
- Baker N.E. and Zitron,A.E. (1995) Drosophila eye development: Notch and Delta amplify a neurogenic pattern conferred on the morphogenic furrow by scabrous. Mech. Dev., 49, 173–189. - PubMed
-
- Baker N.E. and Yu,S.-Y. (1997) Proneural function of neurogenic genes in the developing Drosophila eye. Curr. Biol., 7, 122–132. - PubMed
-
- Baker N.E. and Yu,S.-Y. (1998) The R8-photoreceptor equivalence group in Drosophila: fate choice precedes regulated Delta transcription and is independent of Notch gene dose. Mech. Dev., 74, 3–14. - PubMed
-
- Baker N.E., Mlodzik,M. and Rubin,G.M. (1990) Spacing differentiation in the developing Drosophila eye: a fibrinogen-related lateral inhibitor encodes by scabrous. Science, 250, 1370–1377. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
