

Regulation of cellular differentiation in filamentous cyanobacteria in free-living and plant-associated symbiotic growth states
- PMID: 11875129
- PMCID: PMC120779
- DOI: 10.1128/MMBR.66.1.94-121.2002
Regulation of cellular differentiation in filamentous cyanobacteria in free-living and plant-associated symbiotic growth states
Abstract
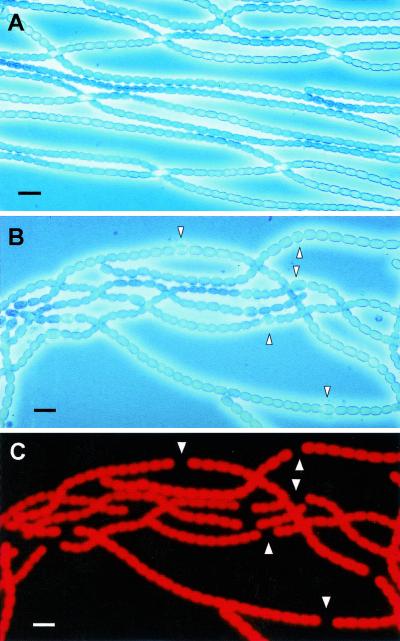
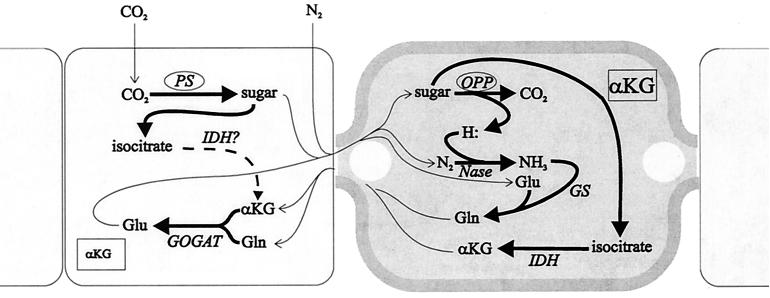
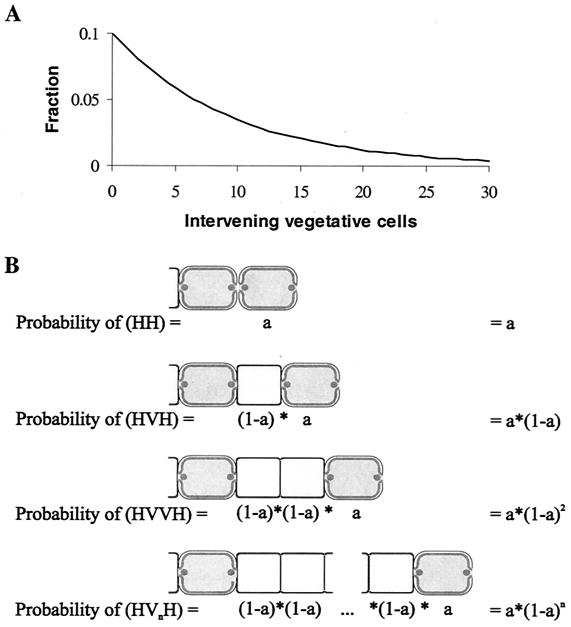
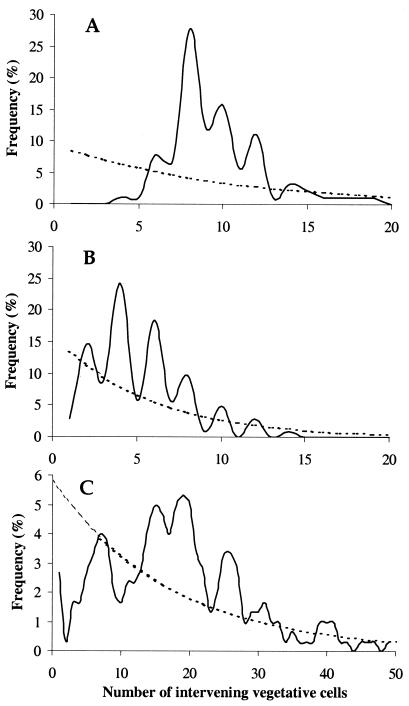
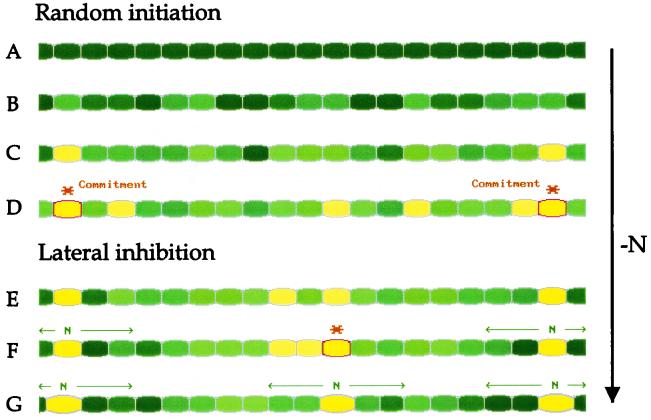
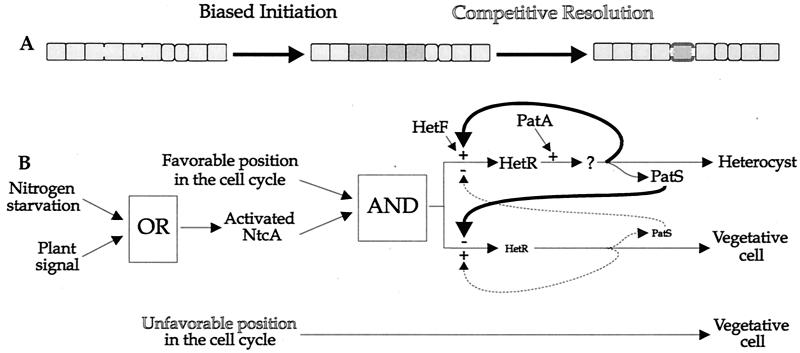
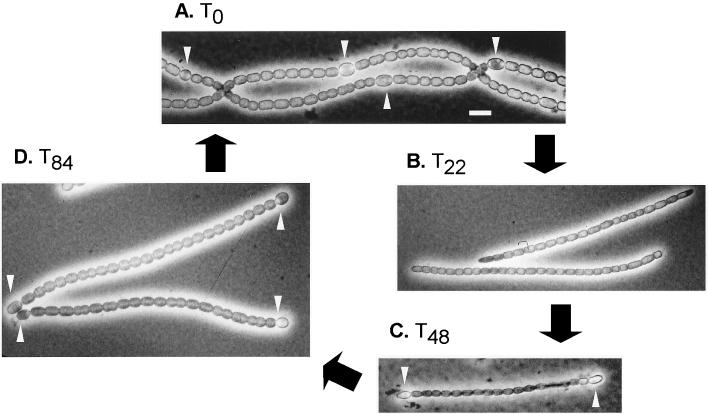
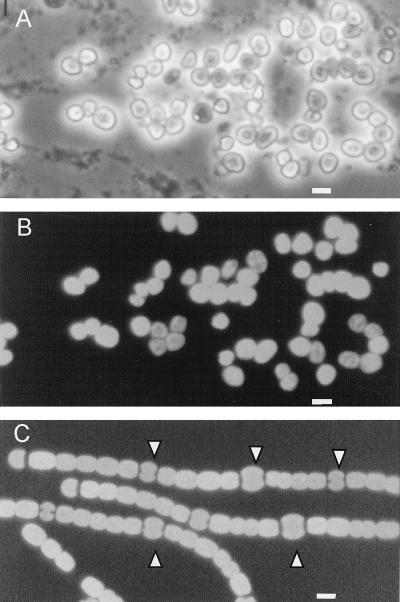
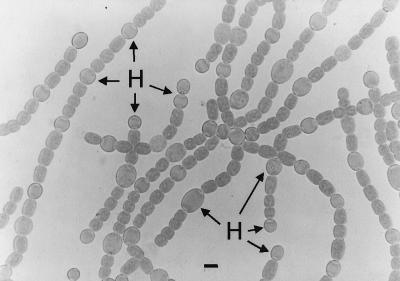
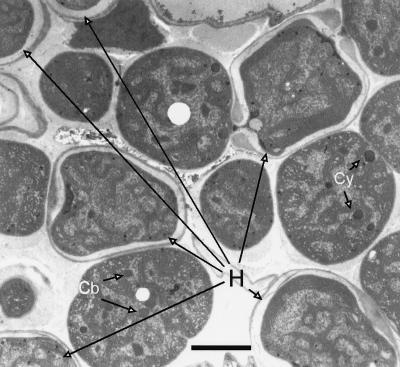
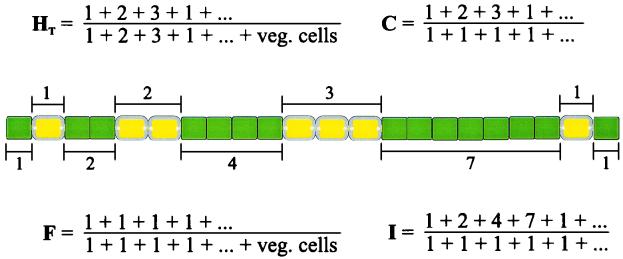
Certain filamentous nitrogen-fixing cyanobacteria generate signals that direct their own multicellular development. They also respond to signals from plants that initiate or modulate differentiation, leading to the establishment of a symbiotic association. An objective of this review is to describe the mechanisms by which free-living cyanobacteria regulate their development and then to consider how plants may exploit cyanobacterial physiology to achieve stable symbioses. Cyanobacteria that are capable of forming plant symbioses can differentiate into motile filaments called hormogonia and into specialized nitrogen-fixing cells called heterocysts. Plant signals exert both positive and negative regulatory control on hormogonium differentiation. Heterocyst differentiation is a highly regulated process, resulting in a regularly spaced pattern of heterocysts in the filament. The evidence is most consistent with the pattern arising in two stages. First, nitrogen limitation triggers a nonrandomly spaced cluster of cells (perhaps at a critical stage of their cell cycle) to initiate differentiation. Interactions between an inhibitory peptide exported by the differentiating cells and an activator protein within them causes one cell within each cluster to fully differentiate, yielding a single mature heterocyst. In symbiosis with plants, heterocyst frequencies are increased 3- to 10-fold because, we propose, either differentiation is initiated at an increased number of sites or resolution of differentiating clusters is incomplete. The physiology of symbiotically associated cyanobacteria raises the prospect that heterocyst differentiation proceeds independently of the nitrogen status of a cell and depends instead on signals produced by the plant partner.
Figures

Similar articles
-
Establishment of a functional symbiosis between the cyanobacterium Nostoc punctiforme and the bryophyte Anthoceros punctatus requires genes involved in nitrogen control and initiation of heterocyst differentiation.Microbiology (Reading). 2002 Jan;148(Pt 1):315-323. doi: 10.1099/00221287-148-1-315. Microbiology (Reading). 2002. PMID: 11782524
-
Cellular differentiation in the cyanobacterium Nostoc punctiforme.Arch Microbiol. 2002 Dec;178(6):395-403. doi: 10.1007/s00203-002-0476-5. Epub 2002 Sep 18. Arch Microbiol. 2002. PMID: 12420158 Review.
-
The Hybrid Histidine Kinase HrmK Is an Early-Acting Factor in the Hormogonium Gene Regulatory Network.J Bacteriol. 2020 Feb 11;202(5):e00675-19. doi: 10.1128/JB.00675-19. Print 2020 Feb 11. J Bacteriol. 2020. PMID: 31792014 Free PMC article.
-
Hormogonium Development and Motility in Filamentous Cyanobacteria.Appl Environ Microbiol. 2023 Jun 28;89(6):e0039223. doi: 10.1128/aem.00392-23. Epub 2023 May 18. Appl Environ Microbiol. 2023. PMID: 37199640 Free PMC article. Review.
-
A Putative O-Linked β-N-Acetylglucosamine Transferase Is Essential for Hormogonium Development and Motility in the Filamentous Cyanobacterium Nostoc punctiforme.J Bacteriol. 2017 Apr 11;199(9):e00075-17. doi: 10.1128/JB.00075-17. Print 2017 May 1. J Bacteriol. 2017. PMID: 28242721 Free PMC article.
Cited by
-
Examining the Evidence for Regulated and Programmed Cell Death in Cyanobacteria. How Significant Are Different Forms of Cell Death in Cyanobacteria Population Dynamics?Front Microbiol. 2021 Mar 22;12:633954. doi: 10.3389/fmicb.2021.633954. eCollection 2021. Front Microbiol. 2021. PMID: 33828539 Free PMC article. Review.
-
Symbiotic Nitrogen Fixation and the Challenges to Its Extension to Nonlegumes.Appl Environ Microbiol. 2016 Jun 13;82(13):3698-3710. doi: 10.1128/AEM.01055-16. Print 2016 Jul 1. Appl Environ Microbiol. 2016. PMID: 27084023 Free PMC article. Review.
-
Perspectives on Endosymbiosis in Coralloid Roots: Association of Cycads and Cyanobacteria.Front Microbiol. 2019 Aug 14;10:1888. doi: 10.3389/fmicb.2019.01888. eCollection 2019. Front Microbiol. 2019. PMID: 31474965 Free PMC article. Review.
-
Nitrogen fixation in eukaryotes--new models for symbiosis.BMC Evol Biol. 2007 Apr 4;7:55. doi: 10.1186/1471-2148-7-55. BMC Evol Biol. 2007. PMID: 17408485 Free PMC article.
-
Mutation at different sites in the Nostoc punctiforme cyaC gene, encoding the multiple-domain enzyme adenylate cyclase, results in different levels of infection of the host plant Blasia pusilla.J Bacteriol. 2008 Mar;190(5):1843-7. doi: 10.1128/JB.01321-07. Epub 2007 Dec 21. J Bacteriol. 2008. PMID: 18156269 Free PMC article.
References
-
- Adams, D. G. 1992. The effect of DL-7-azatryptophan on heterocyst development in the cyanobacterium Anabaena cylindrica. J. Gen. Microbiol. 138:355-362.
-
- Adams, D. G. 2000. Heterocyst formation in cyanobacteria. Curr. Opin. Microbiol. 3:618-624. - PubMed
-
- Adams, D. G. 2000. Symbiotic internactions, p. 523-561. In B. A. Whitton and M. Potts (ed.), The ecology of cyanobacteria. Their diversity in time and space. Kluwer Academic Publishers, Boston, Mass.
-
- Adams, D. G., and N. G. Carr. 1981. Heterocyst differentiation and cell division in the cyanobacterium Anabaena cylindrica: effect of high light intensity. J. Cell Sci. 49:341-352. - PubMed
-
- Adams, D. G., and N. G. Carr. 1989. Control of heterocyst development in the cyanobacterium Anabaena cylindrica. J. Gen. Microbiol. 135:839-849.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
