

Involvement of the cohesin protein, Smc1, in Atm-dependent and independent responses to DNA damage
- PMID: 11877376
- PMCID: PMC155347
- DOI: 10.1101/gad.970602
Involvement of the cohesin protein, Smc1, in Atm-dependent and independent responses to DNA damage
Abstract
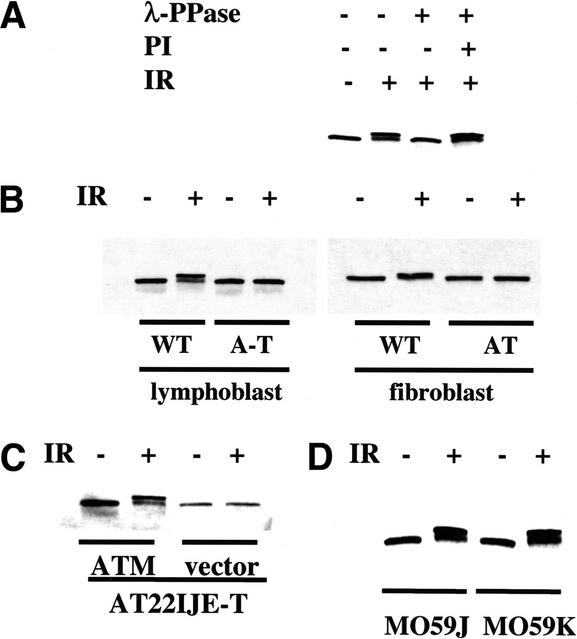
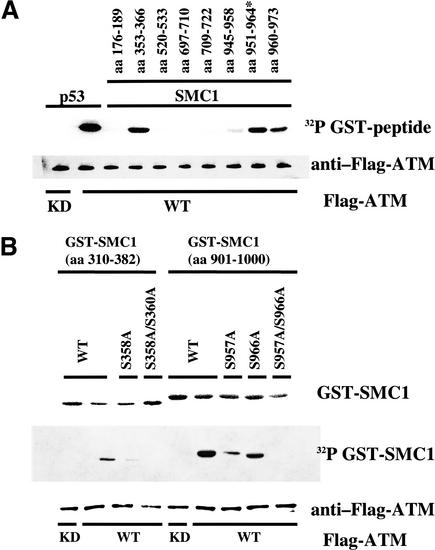
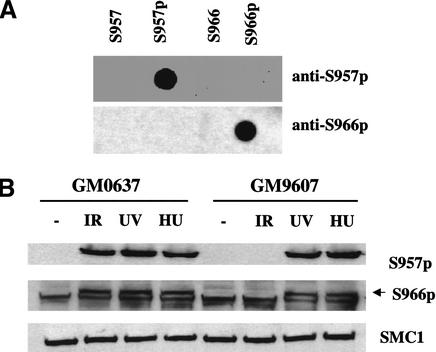
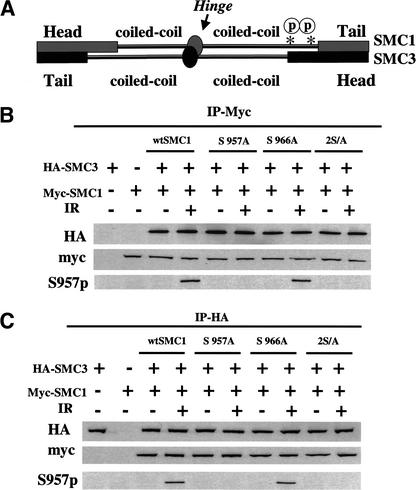
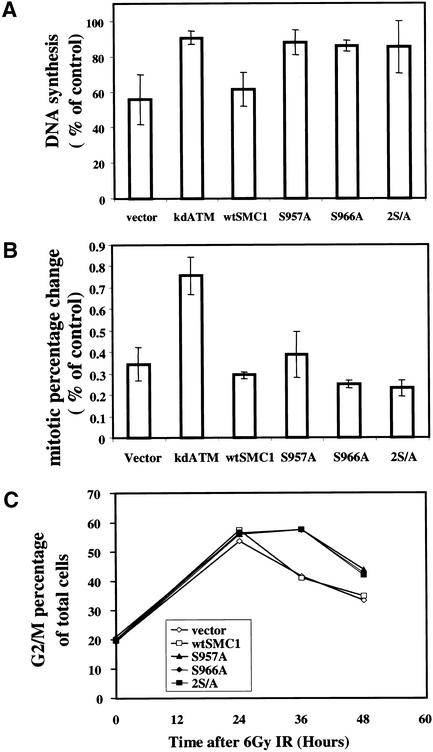
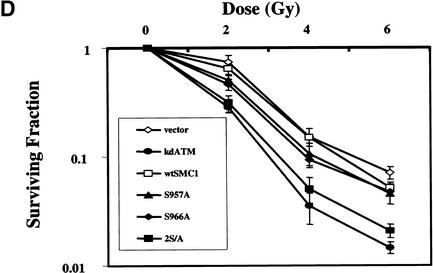
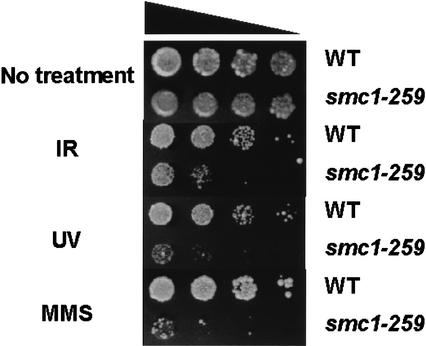
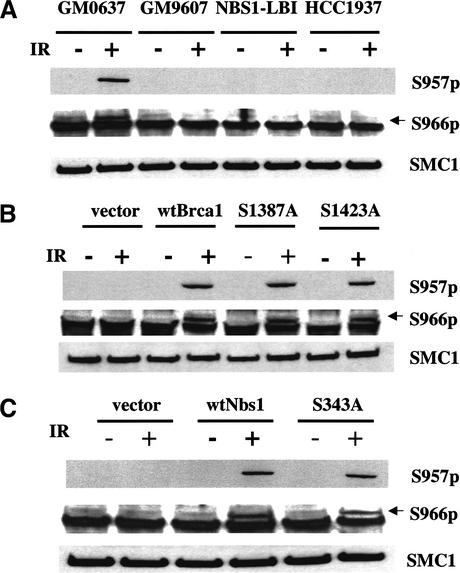
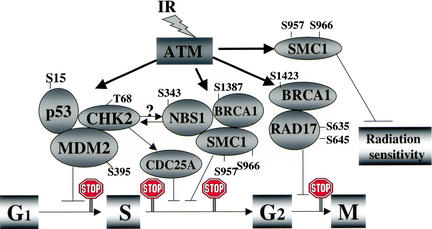
Structural maintenance of chromosomes (SMC) proteins play important roles in sister chromatid cohesion, chromosome condensation, sex-chromosome dosage compensation, and DNA recombination and repair. Protein complexes containing heterodimers of the Smc1 and Smc3 proteins have been implicated specifically in both sister chromatid cohesion and DNA recombination. Here, we show that the protein kinase, Atm, which belongs to a family of phosphatidylinositol 3-kinases that regulate cell cycle checkpoints and DNA recombination and repair, phosphorylates Smc1 protein after ionizing irradiation. Atm phosphorylates Smc1 on serines 957 and 966 in vitro and in vivo, and expression of an Smc1 protein mutated at these phosphorylation sites abrogates the ionizing irradiation-induced S phase cell cycle checkpoint. Optimal phosphorylation of these sites in Smc1 after ionizing irradiation also requires the presence of the Atm substrates Nbs1 and Brca1. These same sites in Smc1 are phosphorylated after treatment with UV irradiation or hydroxyurea in an Atm-independent manner, thus demonstrating that another kinase must be involved in responses to these cellular stresses. Yeast containing hypomorphic mutations in SMC1 and human cells overexpressing Smc1 mutated at both of these phosphorylation sites exhibit decreased survival following ionizing irradiation. These results demonstrate that Smc1 participates in cellular responses to DNA damage and link Smc1 to the Atm signal transduction pathway.
Figures
References
-
- Ahn J-Y, Schwarz JK, Piwnica-Worms H, Canman CE. Threonine 68 phosphorylation by ATM is required for efficient activation of Chk2 in response to ionizing radiation. Cancer Res. 2000;60:5934–5936. - PubMed
-
- Banin S, Moyal L, Shieh S-Y, Taya Y, Anderson CW, Chessa L, Smorodinsky NI, Prives C, Reiss Y, Shiloh Y, et al. Enhanced phosphorylation of p53 by ATM in response to DNA damage. Science. 1998;281:1674–1677. - PubMed
-
- Bao S, Tibbetts RS, Brumbaug KM, Fang Y, Richardson DA, Ali A, Chen SM, Abraham RT, Wang X-F. ATR/ATM-mediated phosphorylation of human Rad17 is required for genotoxic stress responses. Nature. 2001;411:969–974. - PubMed
-
- Canman CE, Lim D-S, Cimprich KA, Taya Y, Tamai K, Sakaguchi K, Appella E, Kastan MB, Siliciano JD. Activation of the ATM kinase by ionizing radiation and phosphorylation of p53. Science. 1998;281:1677–1679. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous