

Lipid raft microdomains: a gateway for compartmentalized trafficking of Ebola and Marburg viruses
- PMID: 11877482
- PMCID: PMC2193767
- DOI: 10.1084/jem.20011500
Lipid raft microdomains: a gateway for compartmentalized trafficking of Ebola and Marburg viruses
Abstract
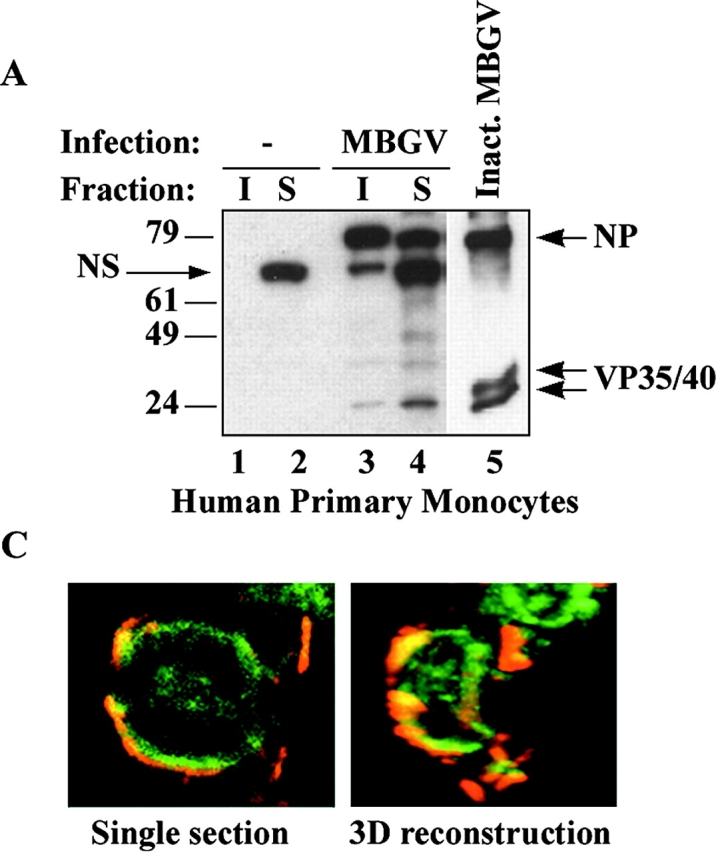
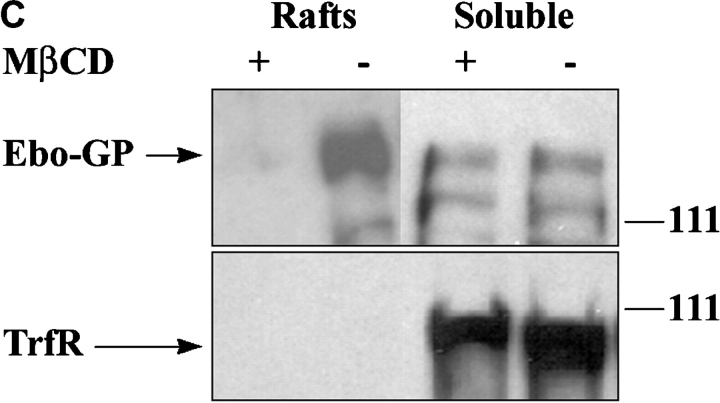
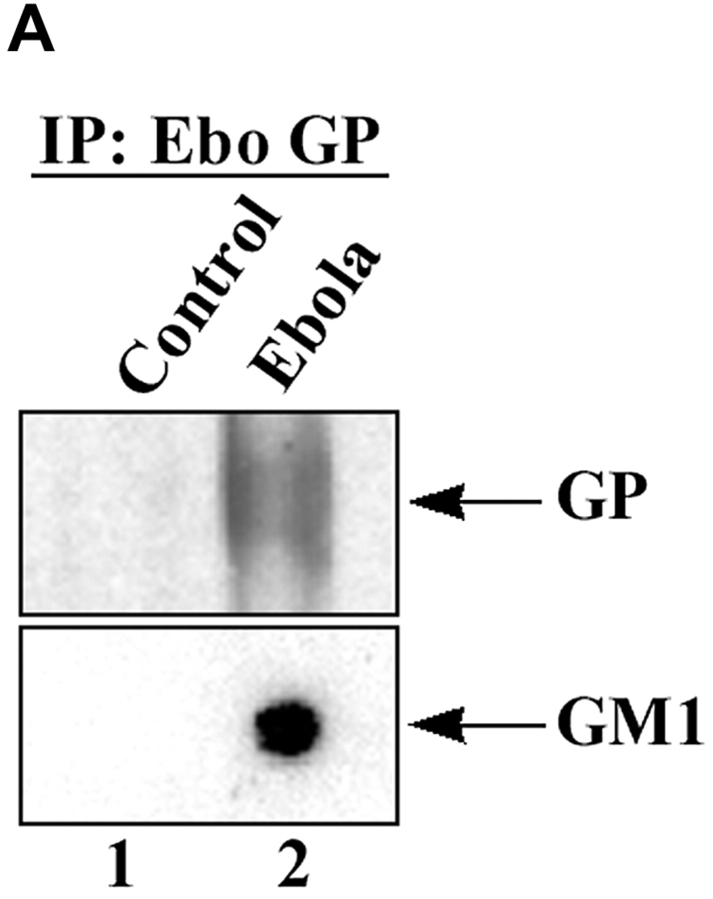
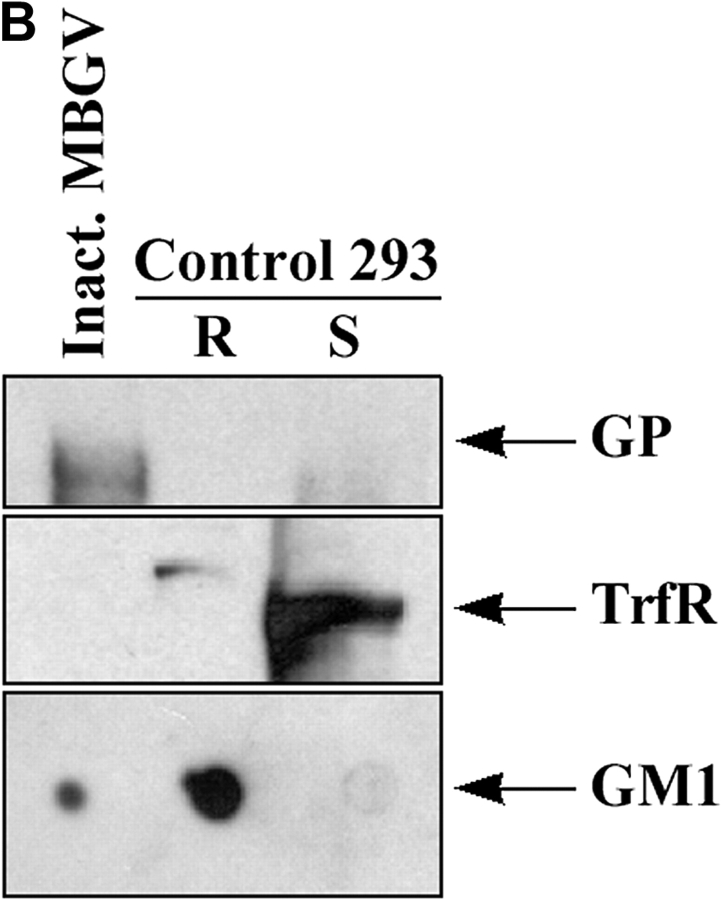
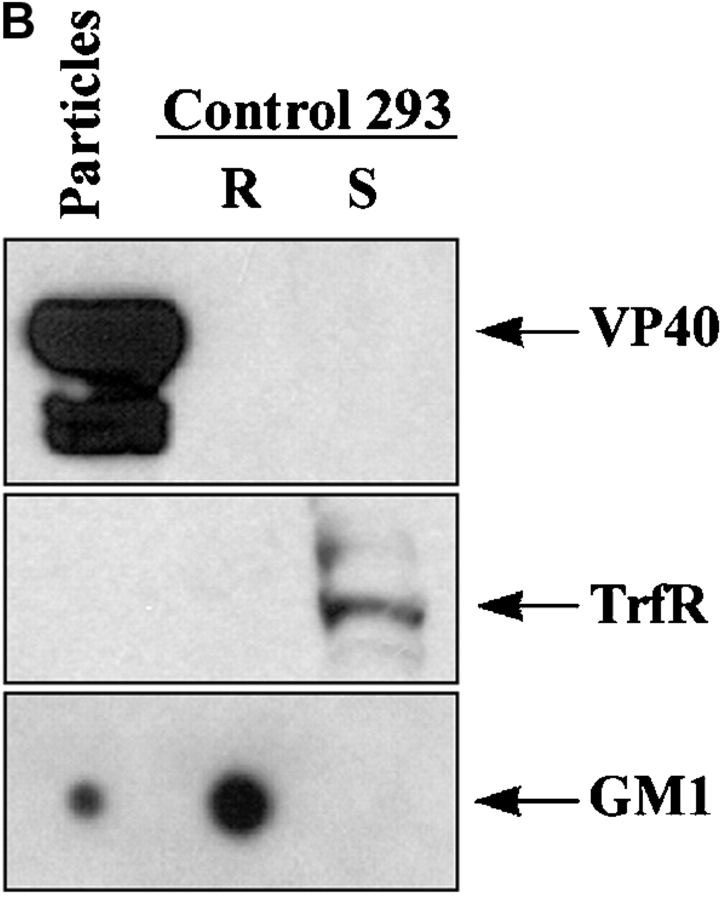
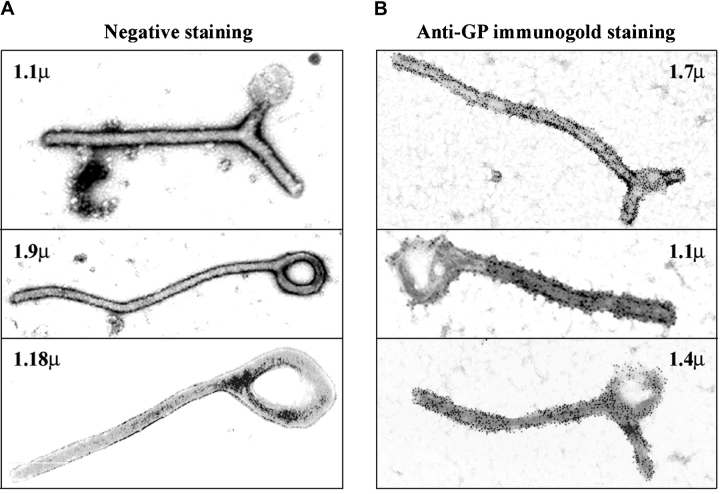
Spatiotemporal aspects of filovirus entry and release are poorly understood. Lipid rafts act as functional platforms for multiple cellular signaling and trafficking processes. Here, we report the compartmentalization of Ebola and Marburg viral proteins within lipid rafts during viral assembly and budding. Filoviruses released from infected cells incorporated raft-associated molecules, suggesting that viral exit occurs at the rafts. Ectopic expression of Ebola matrix protein and glycoprotein supported raft-dependent release of filamentous, virus-like particles (VLPs), strikingly similar to live virus as revealed by electron microscopy. Our findings also revealed that the entry of filoviruses requires functional rafts, identifying rafts as the site of virus attack. The identification of rafts as the gateway for the entry and exit of filoviruses and raft-dependent generation of VLPs have important implications for development of therapeutics and vaccination strategies against infections with Ebola and Marburg viruses.
Figures
References
-
- Feldmann, H., and H.D. Klenk. 1996. Marburg and Ebola viruses. Adv. Virus Res. 47:1–52. - PubMed
-
- Johnson, K.M., J.V. Lange, P.A. Webb, and F.A. Murphy. 1977. Isolation and partial characterisation of a new virus causing acute haemorrhagic fever in Zaire. Lancet. 1:569–571. - PubMed
-
- Jaax, N., P. Jahrling, T. Geisbert, J. Geisbert, K. Steele, K. McKee, D. Nagley, E. Johnson, G. Jaax, and C. Peters. 1995. Transmission of Ebola virus (Zaire strain) to uninfected control monkeys in a biocontainment laboratory. Lancet. 346:1669–1671. - PubMed
-
- Sullivan, N.J., A. Sanchez, P.E. Rollin, Z.Y. Yang, and G.J. Nabel. 2000. Development of a preventive vaccine for Ebola virus infection in primates. Nature. 408:605–609. - PubMed
-
- Vanderzanden, L., M. Bray, D. Fuller, T. Roberts, D. Custer, K. Spik, P. Jahrling, J. Huggins, A. Schmaljohn, and C. Schmaljohn. 1998. DNA vaccines expressing either the GP or NP genes of Ebola virus protect mice from lethal challenge. Virology. 246:134–144. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
