

Natural killer T cell ligand alpha-galactosylceramide enhances protective immunity induced by malaria vaccines
- PMID: 11877484
- PMCID: PMC2193764
- DOI: 10.1084/jem.20011889
Natural killer T cell ligand alpha-galactosylceramide enhances protective immunity induced by malaria vaccines
Abstract
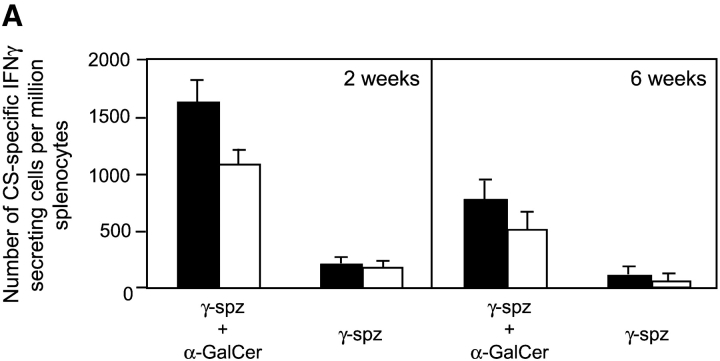
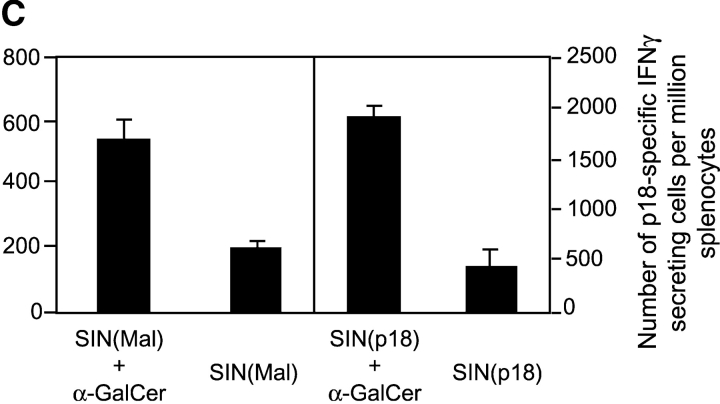
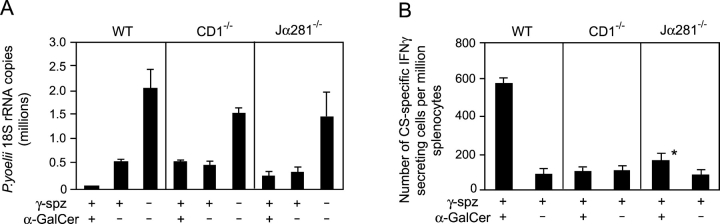
The important role played by CD8(+) T lymphocytes in the control of parasitic and viral infections, as well as tumor development, has raised the need for the development of adjuvants capable of enhancing cell-mediated immunity. It is well established that protective immunity against liver stages of malaria parasites is primarily mediated by CD8(+) T cells in mice. Activation of natural killer T (NKT) cells by the glycolipid ligand, alpha-galactosylceramide (alpha-GalCer), causes bystander activation of NK, B, CD4(+), and CD8(+) T cells. Our study shows that coadministration of alpha-GalCer with suboptimal doses of irradiated sporozoites or recombinant viruses expressing a malaria antigen greatly enhances the level of protective anti-malaria immunity in mice. We also show that coadministration of alpha-GalCer with various different immunogens strongly enhances antigen-specific CD8(+) T cell responses, and to a lesser degree, Th1-type responses. The adjuvant effects of alpha-GalCer require CD1d molecules, Valpha14 NKT cells, and interferon gamma. As alpha-GalCer stimulates both human and murine NKT cells, these findings should contribute to the design of more effective vaccines against malaria and other intracellular pathogens, as well as tumors.
Figures






Comment in
-
Adjuvants of immunity: harnessing innate immunity to promote adaptive immunity.J Exp Med. 2002 Mar 4;195(5):F19-23. doi: 10.1084/jem.20020073. J Exp Med. 2002. PMID: 11877490 Free PMC article. No abstract available.
References
-
- Seder, R.A., and A.V. Hill. 2000. Vaccines against intracellular infections requiring cellular immunity. Nature. 406:793–798. - PubMed
-
- Smyth, M.J., D.T. Godfrey, and J.A. Trapani. 2001. A fresh look at tumor immunosurveillance and immunotherapy. Nat. Immunol. 2:293–299. - PubMed
-
- Pardoll, D.M., and S.L. Topalian. 1998. The role of CD4+ T cell responses in antitumor immunity. Curr. Opin. Immunol. 10:588–594. - PubMed
-
- Singh, M., and D. O'Hagan. 1999. Advances in vaccine adjuvants. Nat. Biotechnol. 17:1075–1081. - PubMed
-
- de Gruijl, T.D., and D.T. Curiel. 1999. Cancer vaccine strategies get bigger and better. Nat. Med. 5:1124–1125. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
