

Crystal structure of the yeast cytochrome bc1 complex with its bound substrate cytochrome c
- PMID: 11880631
- PMCID: PMC122428
- DOI: 10.1073/pnas.052704699
Crystal structure of the yeast cytochrome bc1 complex with its bound substrate cytochrome c
Abstract
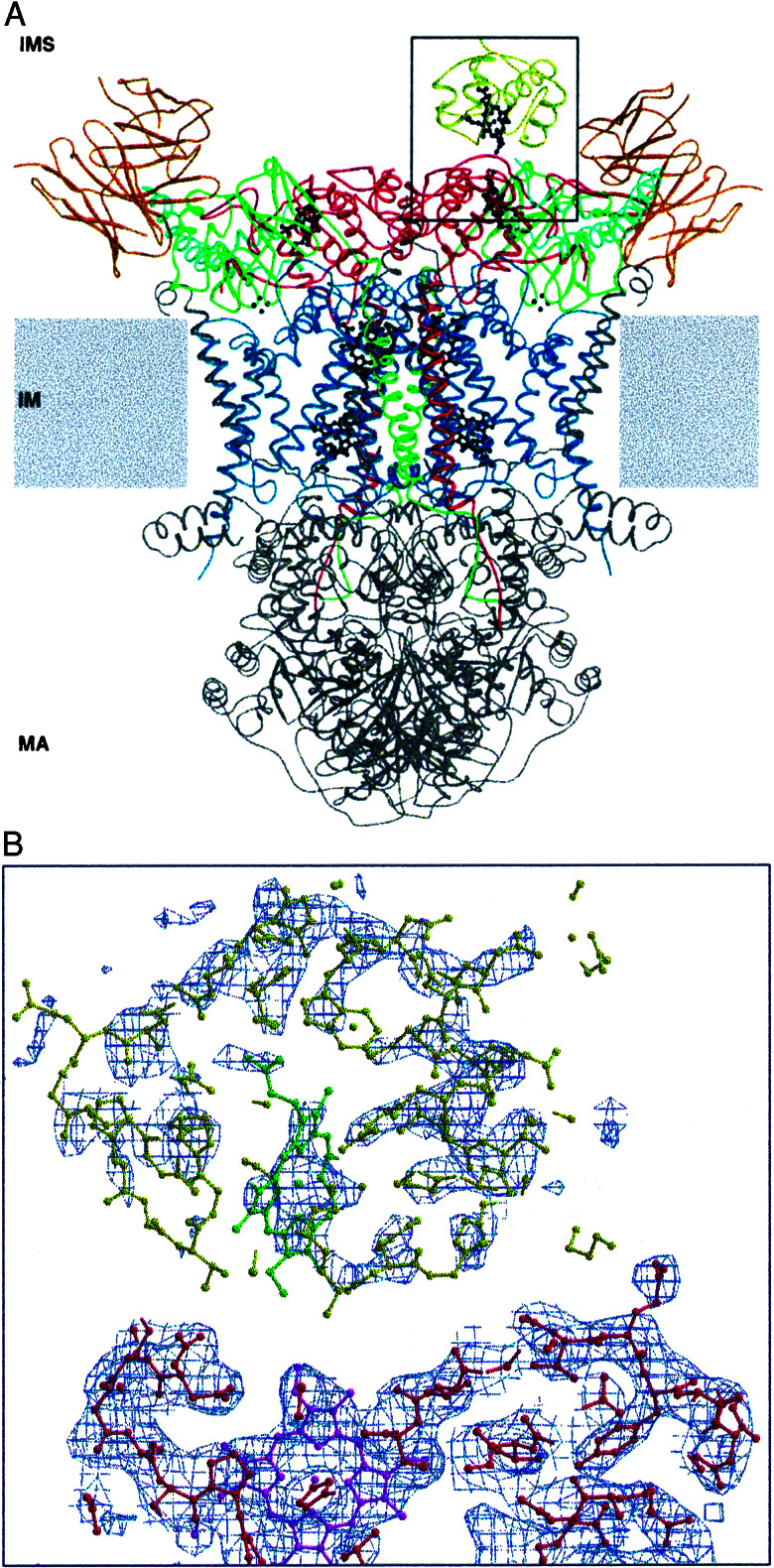
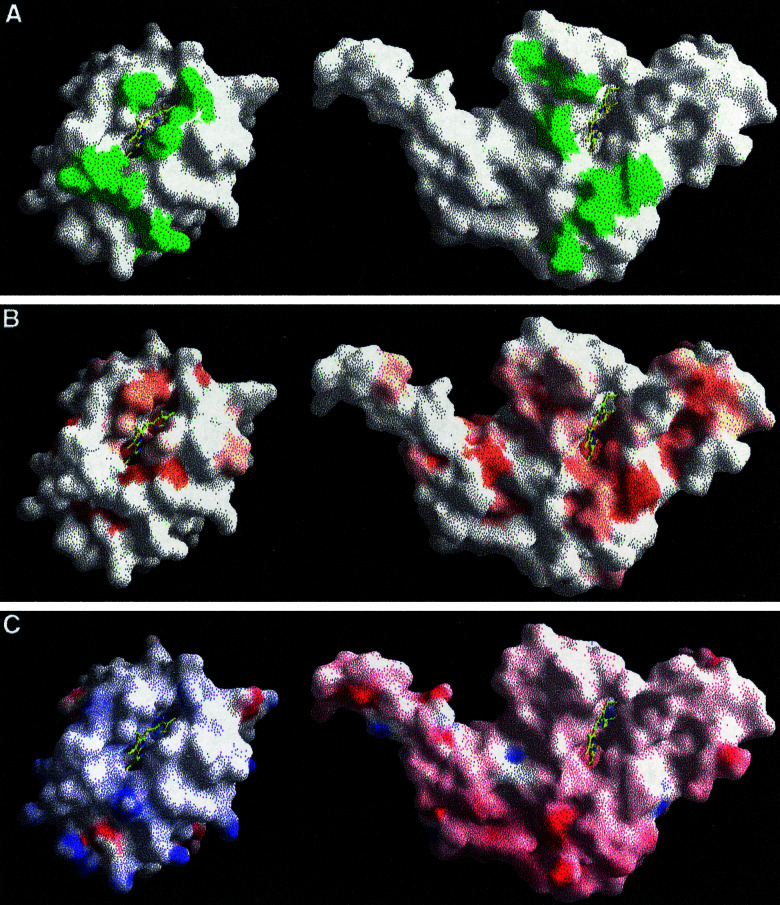
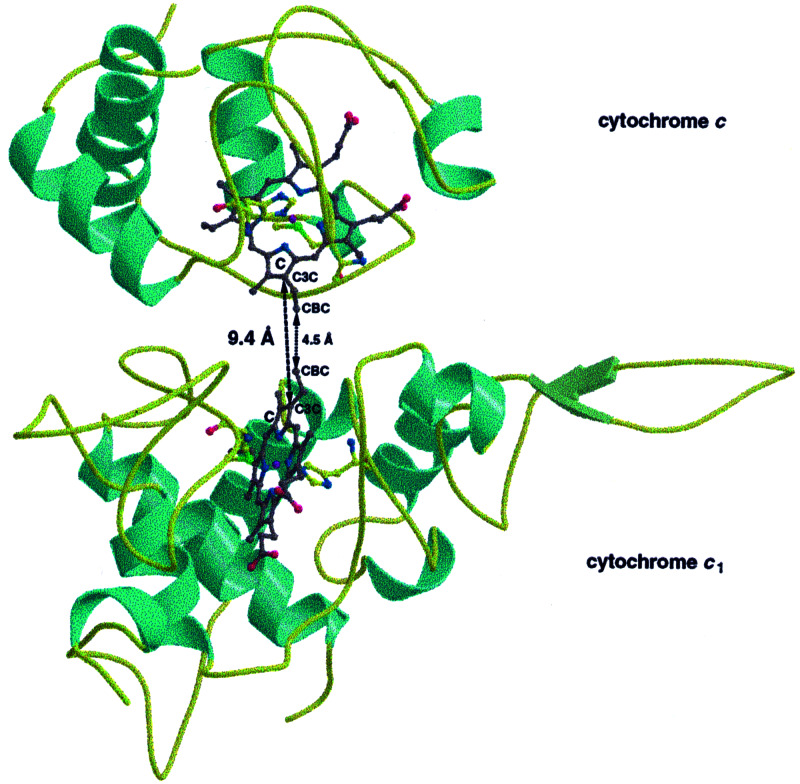
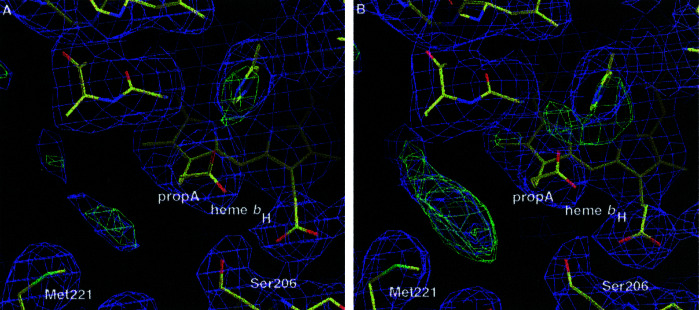
Small diffusible redox proteins facilitate electron transfer in respiration and photosynthesis by alternately binding to integral membrane proteins. Specific and transient complexes need to be formed between the redox partners to ensure fast turnover. In respiration, the mobile electron carrier cytochrome c shuttles electrons from the cytochrome bc1 complex to cytochrome c oxidase. Despite extensive studies of this fundamental step of energy metabolism, the structures of the respective electron transfer complexes were not known. Here we present the crystal structure of the complex between cytochrome c and the cytochrome bc1 complex from Saccharomyces cerevisiae. The complex was crystallized with the help of an antibody fragment, and its structure was determined at 2.97-A resolution. Cytochrome c is bound to subunit cytochrome c1 of the enzyme. The tight and specific interactions critical for electron transfer are mediated mainly by nonpolar forces. The close spatial arrangement of the c-type hemes unexpectedly suggests a direct and rapid heme-to-heme electron transfer at a calculated rate of up to 8.3 x 10(6) s(-1). Remarkably, cytochrome c binds to only one recognition site of the homodimeric multisubunit complex. Interestingly, the occupancy of quinone in the Qi site is higher in the monomer with bound cytochrome c, suggesting a coordinated binding and reduction of both electron-accepting substrates. Obviously, cytochrome c reduction by the cytochrome bc1 complex can be regulated in response to respiratory conditions.
Figures
References
-
- Saraste M. Science. 1999;283:1488–1493. - PubMed
-
- Mitchell P. J Theor Biol. 1976;62:327–367. - PubMed
-
- Brandt U, Trumpower B L. Crit Rev Biochem Mol Biol. 1994;29:165–197. - PubMed
-
- Yu C-A, Yu L, King T E. J Biol Chem. 1973;248:528–533. - PubMed
-
- Speck S H, Margoliash E. J Biol Chem. 1984;259:1064–1072. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
