

Muscle contraction under capillaries in hamster muscle induces arteriolar dilatation via K(ATP) channels and nitric oxide
- PMID: 11882686
- PMCID: PMC2290146
- DOI: 10.1113/jphysiol.2001.013388
Muscle contraction under capillaries in hamster muscle induces arteriolar dilatation via K(ATP) channels and nitric oxide
Abstract
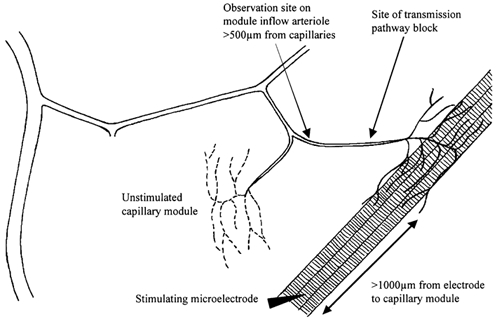
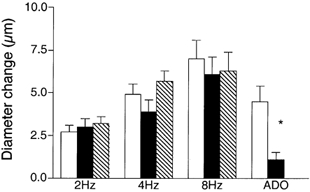
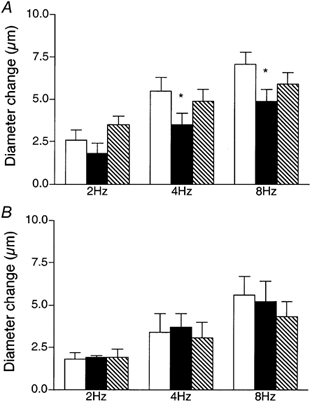
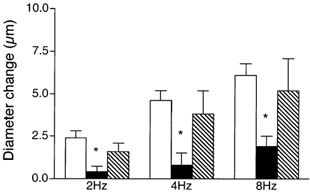
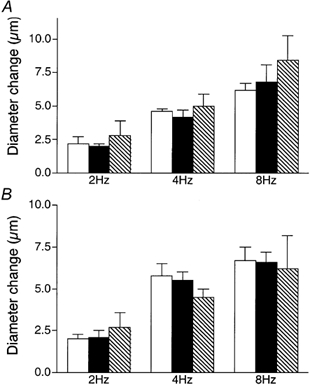
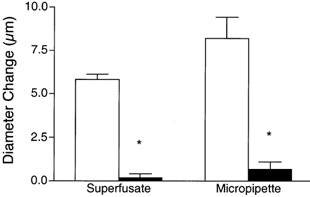
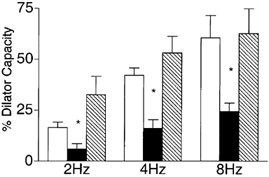
We tested the hypothesis that adenosine and nitric oxide can be sensed by capillaries and are implicated in the remote arteriolar dilatation initiated by muscle contraction. We also explored a role for K(ATP) channel activity in this response. Small bundles of muscle fibres underlying a group of capillaries in cremaster muscles of anaesthetized hamsters were electrically stimulated to contract for 2 min at each of 2, 4 and 8 Hz. Diameter changes were measured in the inflow arteriole to the group of capillaries after muscle contraction in the presence or absence of 10(-6) M xanthine amine congener (XAC) to block A(1) and A(2) adenosine receptors, 10(-4) or 10(-3) M N(omega)-nitro-L-arginine (LNNA) to block nitric oxide production, or 10(-5) M glibenclamide to block K(ATP) channel activity. Dilatations were unchanged with XAC (3.0 +/- 0.5, 3.9 +/- 0.7 and 6.1 +/- 1.0 microm), but were significantly reduced with LNNA (to 1.8 +/- 0.6, 3.5 +/- 0.7 and 4.9 +/- 0.7 microm) or glibenclamide (to 0.4 +/- 0.3, 0.8 +/- 0.7 and 1.9 +/- 0.6 microm). Neither K(ATP) channel activity nor nitric oxide was required for transmission or manifestation of the dilator response. Thus, muscle contraction can be sensed by capillaries and the signalling mechanism for the ensuing remote dilatation depends on K(ATP) channel activity and on NO, but not adenosine. Local application of 10(-4) M adenosine, 10(-4) M sodium nitroprusside or 10(-5) M pinacidil directly to capillaries initiated remote arteriolar dilatations. Thus, capillaries can respond directly to known mediators of metabolic vasodilatation, but these signalling pathways are not invariably implicated in the response to muscle contraction.
Figures
References
-
- Baez S. An open cremaster preparationfor the study of blood vessels by in vivo microscopy. Microvascular Research. 1973;5:384–394. - PubMed
-
- Berg BR, Cohen KD, Sarelius IH. Direct coupling between blood flow and metabolism at the capillary level in striated muscle. American Journal of Physiology. 1997;272:H2693–2700. - PubMed
-
- Berg BR, Sarelius IH. Functional capillary organization in striated muscle. American Journal of Physiology. 1995;268:H1215–1222. - PubMed
-
- Brayden JE. Membrane hyperpolarization is a mechanism of endothelium-dependent cerebral vasodilation. American Journal of Physiology. 1990;259:H668–673. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources