

Different temporal processing of sensory inputs in the rat thalamus during quiescent and information processing states in vivo
- PMID: 11882688
- PMCID: PMC2290158
- DOI: 10.1113/jphysiol.2001.013283
Different temporal processing of sensory inputs in the rat thalamus during quiescent and information processing states in vivo
Abstract
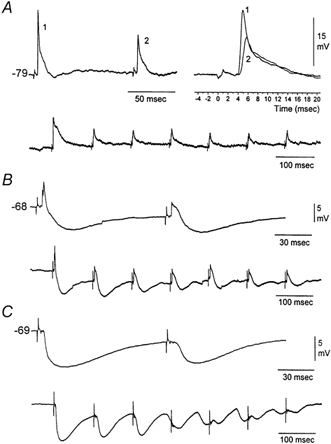
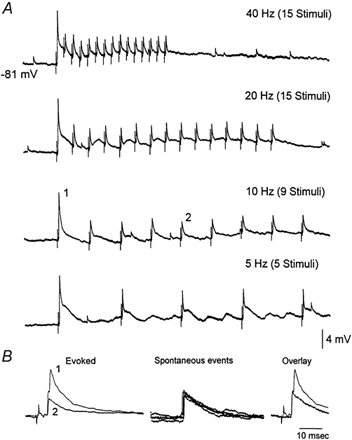
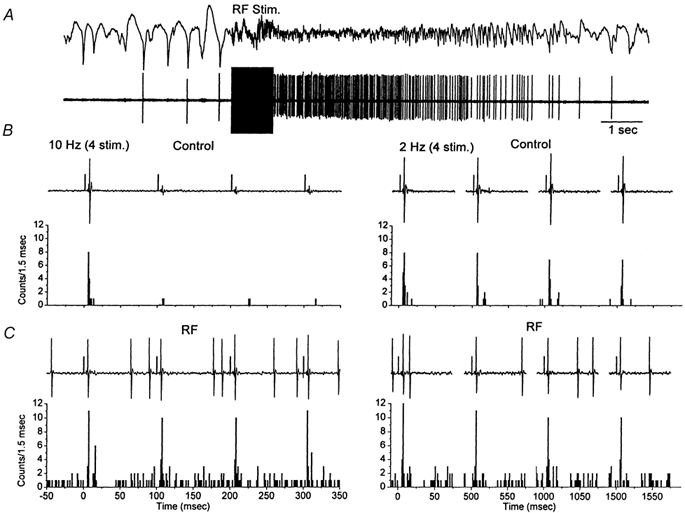
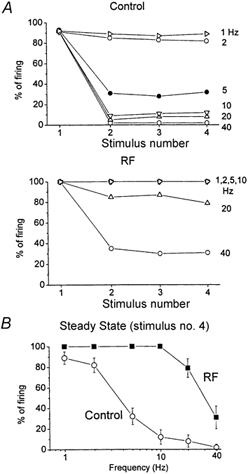
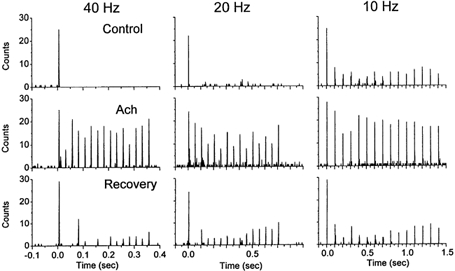
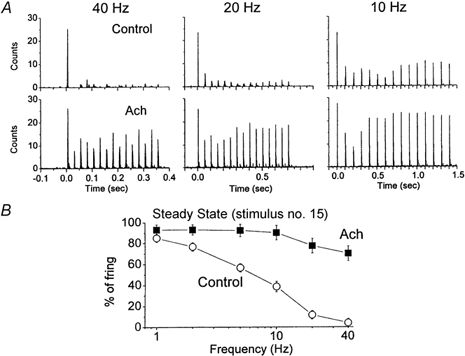
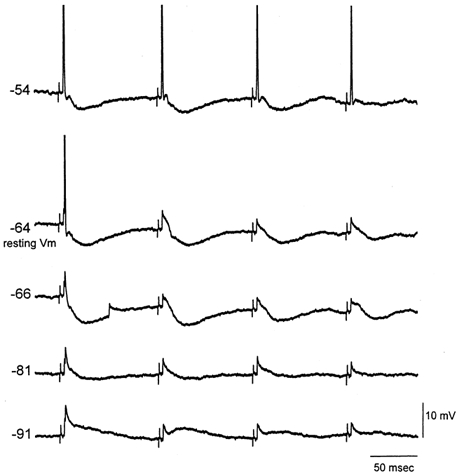
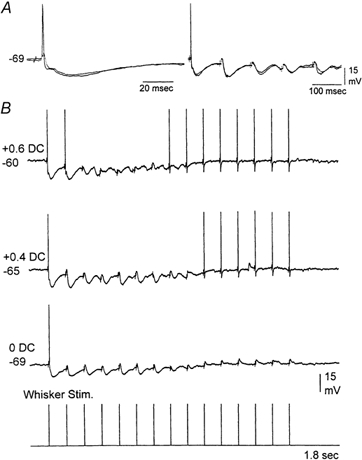
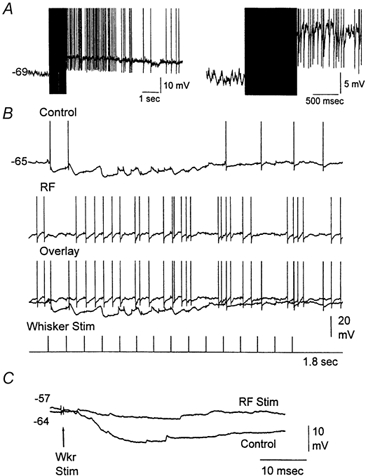
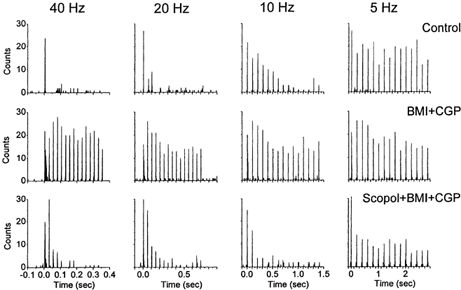
Sensory inputs from the whiskers reach the primary somatosensory thalamus through the medial lemniscus tract. The main role of the thalamus is to relay these sensory inputs to the neocortex according to the regulations dictated by behavioural state. Intracellular recordings in urethane-anaesthetized rats show that whisker stimulation evokes EPSP-IPSP sequences in thalamic neurons. Both EPSPs and IPSPs depress with repetitive whisker stimulation at frequencies above 2 Hz. Single-unit recordings reveal that during quiescent states thalamic responses to repetitive whisker stimulation are suppressed at frequencies above 2 Hz, so that only low-frequency sensory stimulation is relayed to the neocortex. In contrast, during activated states, induced by stimulation of the brainstem reticular formation or application of acetylcholine in the thalamus, high-frequency whisker stimulation at up to 40 Hz is relayed to the neocortex. Sensory suppression is caused by the depression of lemniscal EPSPs in relatively hyperpolarized thalamocortical neurons. Sensory suppression is abolished during activated states because thalamocortical neurons depolarize and the depressed lemniscal EPSPs are able to reach firing threshold. Strong IPSPs may also contribute to sensory suppression by hyperpolarizing thalamocortical neurons, but during activated states IPSPs are strongly reduced altogether. The results indicate that the synaptic depression of lemniscal EPSPs and the level of depolarization of thalamocortical neurons work together in thalamic primary sensory pathways to suppress high-frequency sensory inputs during non-activated (quiescent) states while permitting the faithful relay of high-frequency sensory information during activated (processing) states.
Figures
Similar articles
-
Properties of primary sensory (lemniscal) synapses in the ventrobasal thalamus and the relay of high-frequency sensory inputs.J Neurophysiol. 2002 Feb;87(2):946-53. doi: 10.1152/jn.00426.2001. J Neurophysiol. 2002. PMID: 11826059
-
Role of thalamocortical sensory suppression during arousal: focusing sensory inputs in neocortex.J Neurosci. 2002 Nov 15;22(22):9651-5. doi: 10.1523/JNEUROSCI.22-22-09651.2002. J Neurosci. 2002. PMID: 12427819 Free PMC article.
-
Spatiotemporal gating of sensory inputs in thalamus during quiescent and activated states.J Neurosci. 2005 Nov 23;25(47):10990-1002. doi: 10.1523/JNEUROSCI.3229-05.2005. J Neurosci. 2005. PMID: 16306412 Free PMC article.
-
Dynamics of sensory thalamocortical synaptic networks during information processing states.Prog Neurobiol. 2004 Nov;74(4):213-47. doi: 10.1016/j.pneurobio.2004.09.002. Prog Neurobiol. 2004. PMID: 15556288 Review.
-
Map transfer from the thalamus to the neocortex: inputs from the barrel field.Semin Cell Dev Biol. 2014 Nov;35:147-55. doi: 10.1016/j.semcdb.2014.07.005. Epub 2014 Jul 11. Semin Cell Dev Biol. 2014. PMID: 25020201 Review.
Cited by
-
Imbalance between excitation and inhibition in the somatosensory cortex produces postadaptation facilitation.J Neurosci. 2013 May 8;33(19):8463-71. doi: 10.1523/JNEUROSCI.4845-12.2013. J Neurosci. 2013. PMID: 23658183 Free PMC article.
-
Coding of apparent motion in the thalamic nucleus of the rat vibrissal somatosensory system.J Neurosci. 2012 Mar 7;32(10):3339-51. doi: 10.1523/JNEUROSCI.3890-11.2012. J Neurosci. 2012. PMID: 22399756 Free PMC article.
-
Thalamocortical Up states: differential effects of intrinsic and extrinsic cortical inputs on persistent activity.J Neurosci. 2007 Apr 18;27(16):4261-72. doi: 10.1523/JNEUROSCI.0003-07.2007. J Neurosci. 2007. PMID: 17442810 Free PMC article.
-
Cortex dynamically modulates responses of thalamic relay neurons through prolonged circuit-level disinhibition in rat thalamus in vivo.J Neurophysiol. 2016 Nov 1;116(5):2368-2382. doi: 10.1152/jn.00424.2016. Epub 2016 Aug 31. J Neurophysiol. 2016. PMID: 27582292 Free PMC article.
-
A narrative on the neurobiological roots of attachment-system functioning.Commun Psychol. 2024 Oct 16;2(1):96. doi: 10.1038/s44271-024-00147-9. Commun Psychol. 2024. PMID: 39406946 Free PMC article. Review.
References
-
- Ahissar E, Sosnik R, Haidarliu S. Transformation from temporal to rate coding in a somatosensory thalamocortical pathway. Nature. 2000;406:302–306. - PubMed
-
- Andersen P, Andersson SA, Landgren S. Some properties of the thalamic relay cells in the spino-cervico-lemniscal path. Acta Physiologica Scandinavica. 1966;68:72–83.
-
- Armstrong-James M, Callahan CA. Thalamo-cortical processing of vibrissal information in the rat. II. spatiotemporal convergence in the thalamic ventroposterior medial nucleus (VPm) and its relevance to generation of receptive fields of S1 cortical ‘barrel’ neurones. Journal of Comparative Neurology. 1991;303:211–224. - PubMed
-
- Baldissera F, Margnelli M. Disynaptic IPSPs of lemniscal origin in relay-cells of thalamic ventrobasal nuclear complex of the cat. Brain Research. 1979;163:171–175. - PubMed
-
- Barbaresi P, Spreafico R, Frassoni C, Rustioni A. GABAergic neurons are present in the dorsal column nuclei but not in the ventroposterior complex of rats. Brain Research. 1986;382:305–326. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources