

Novel swine virulence determinant in the left variable region of the African swine fever virus genome
- PMID: 11884534
- PMCID: PMC136047
- DOI: 10.1128/jvi.76.7.3095-3104.2002
Novel swine virulence determinant in the left variable region of the African swine fever virus genome
Abstract
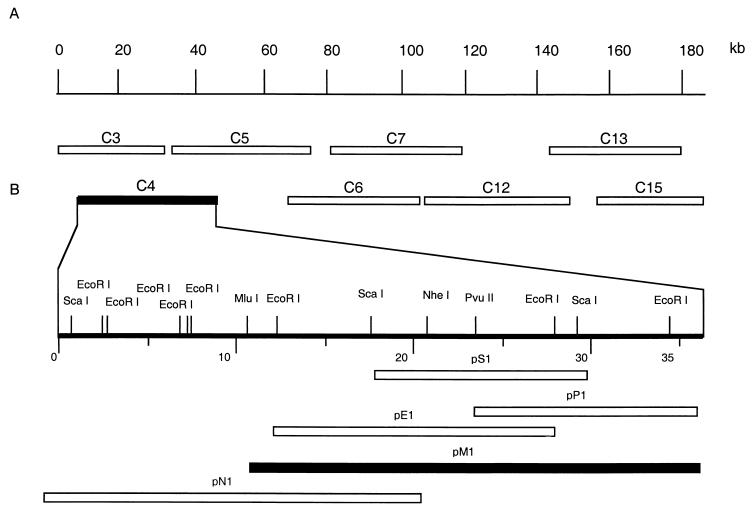
Previously we have shown that the African swine fever virus (ASFV) NL gene deletion mutant E70DeltaNL is attenuated in pigs. Our recent observations that NL gene deletion mutants of two additional pathogenic ASFV isolates, Malawi Lil-20/1 and Pr4, remained highly virulent in swine (100% mortality) suggested that these isolates encoded an additional virulence determinant(s) that was absent from E70. To map this putative virulence determinant, in vivo marker rescue experiments were performed by inoculating swine with infection-transfection lysates containing E70 NL deletion mutant virus (E70DeltaNL) and cosmid DNA clones from the Malawi NL gene deletion mutant (MalDeltaNL). A cosmid clone representing the left-hand 38-kb region (map units 0.05 to 0.26) of the MalDeltaNL genome was capable of restoring full virulence to E70DeltaNL. Southern blot analysis of recovered virulent viruses confirmed that they were recombinant E70DeltaNL genomes containing a 23- to 28-kb DNA fragment of the Malawi genome. These recombinants exhibited an unaltered MalDeltaNL disease and virulence phenotype when inoculated into swine. Additional in vivo marker rescue experiments identified a 20-kb fragment, encoding members of multigene families (MGF) 360 and 530, as being capable of fully restoring virulence to E70DeltaNL. Comparative nucleotide sequence analysis of the left variable region of the E70DeltaNL and Malawi Lil-20/1 genomes identified an 8-kb deletion in the E70DeltaNL isolate which resulted in the deletion and/or truncation of three MGF 360 genes and four MGF 530 genes. A recombinant MalDeltaNL deletion mutant lacking three members of each MGF gene family was constructed and evaluated for virulence in swine. The mutant virus replicated normally in macrophage cell culture but was avirulent in swine. Together, these results indicate that a region within the left variable region of the ASFV genome containing the MGF 360 and 530 genes represents a previously unrecognized virulence determinant for domestic swine.
Figures
Similar articles
-
The first complete genome sequence of the African swine fever virus genotype X and serogroup 7 isolated in domestic pigs from the Democratic Republic of Congo.Virol J. 2021 Jan 21;18(1):23. doi: 10.1186/s12985-021-01497-0. Virol J. 2021. PMID: 33478547 Free PMC article.
-
An African swine fever virus virulence-associated gene NL-S with similarity to the herpes simplex virus ICP34.5 gene.J Virol. 1996 Dec;70(12):8865-71. doi: 10.1128/JVI.70.12.8865-8871.1996. J Virol. 1996. PMID: 8971015 Free PMC article.
-
A BIR motif containing gene of African swine fever virus, 4CL, is nonessential for growth in vitro and viral virulence.Virology. 1997 Apr 14;230(2):252-64. doi: 10.1006/viro.1997.8481. Virology. 1997. PMID: 9143281
-
Differential Effect of Deleting Members of African Swine Fever Virus Multigene Families 360 and 505 from the Genotype II Georgia 2007/1 Isolate on Virus Replication, Virulence, and Induction of Protection.J Virol. 2022 Mar 23;96(6):e0189921. doi: 10.1128/jvi.01899-21. Epub 2022 Jan 19. J Virol. 2022. PMID: 35044212 Free PMC article.
-
African swine fever virus NL gene is not required for virus virulence.J Gen Virol. 1998 Oct;79 ( Pt 10):2543-7. doi: 10.1099/0022-1317-79-10-2543. J Gen Virol. 1998. PMID: 9780062
Cited by
-
Inhibition of a large double-stranded DNA virus by MxA protein.J Virol. 2009 Mar;83(5):2310-20. doi: 10.1128/JVI.00781-08. Epub 2008 Dec 24. J Virol. 2009. PMID: 19109387 Free PMC article.
-
The first complete genome sequence of the African swine fever virus genotype X and serogroup 7 isolated in domestic pigs from the Democratic Republic of Congo.Virol J. 2021 Jan 21;18(1):23. doi: 10.1186/s12985-021-01497-0. Virol J. 2021. PMID: 33478547 Free PMC article.
-
Development and evaluation of duplex TaqMan real-time PCR assay for detection and differentiation of wide-type and MGF505-2R gene-deleted African swine fever viruses.BMC Vet Res. 2020 Nov 9;16(1):428. doi: 10.1186/s12917-020-02639-2. BMC Vet Res. 2020. PMID: 33167979 Free PMC article.
-
African swine fever virus multigene family 360 and 530 genes affect host interferon response.J Virol. 2004 Feb;78(4):1858-64. doi: 10.1128/jvi.78.4.1858-1864.2004. J Virol. 2004. PMID: 14747550 Free PMC article.
-
African Swine Fever Virus and Host Response: Transcriptome Profiling of the Georgia 2007/1 Strain and Porcine Macrophages.J Virol. 2022 Mar 9;96(5):e0193921. doi: 10.1128/jvi.01939-21. Epub 2022 Jan 12. J Virol. 2022. PMID: 35019713 Free PMC article.
References
-
- Afonso, C. L., L. Zsak, C. Carrillo, M. V. Borca, and D. L. Rock. 1998. African swine fever virus NL gene is not required for virus virulence. J. Gen. Virol. 79:2543-2547. - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
