

The fusion peptide of Semliki Forest virus associates with sterol-rich membrane domains
- PMID: 11884551
- PMCID: PMC136022
- DOI: 10.1128/jvi.76.7.3267-3275.2002
The fusion peptide of Semliki Forest virus associates with sterol-rich membrane domains
Abstract
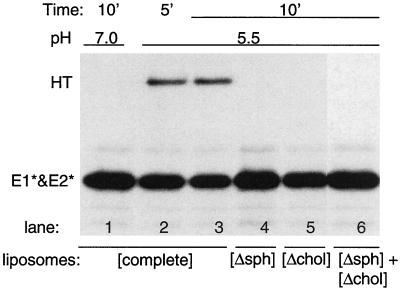
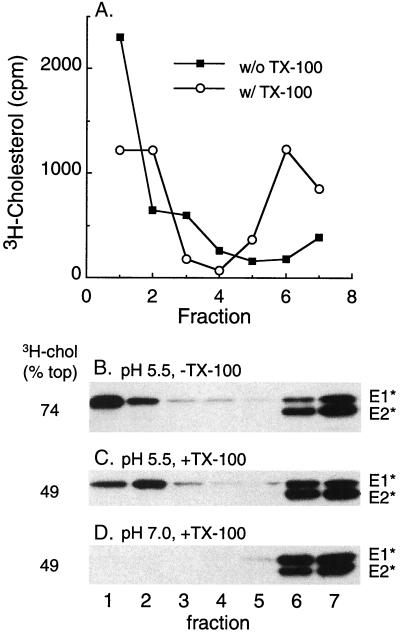
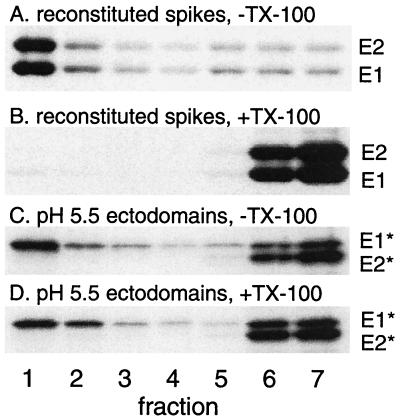
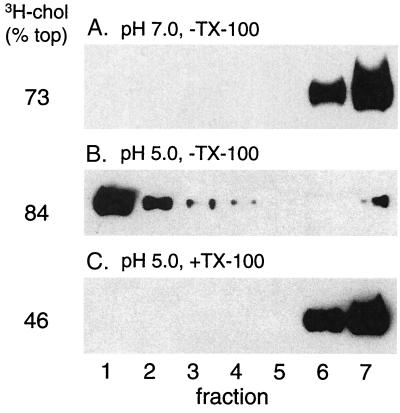
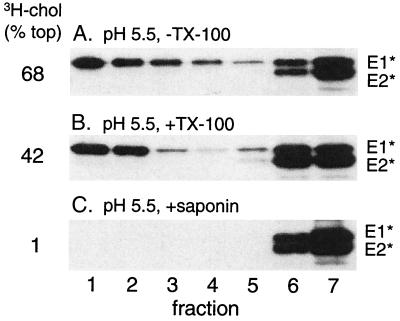
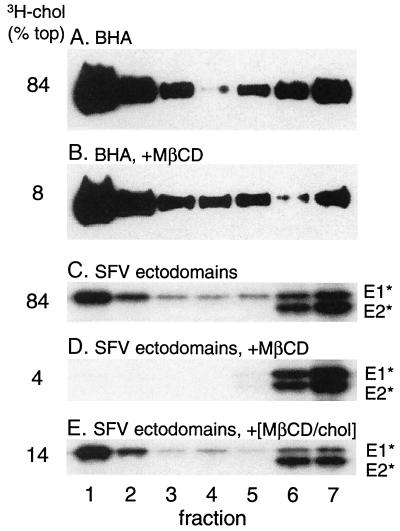
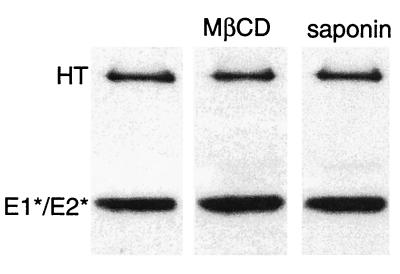
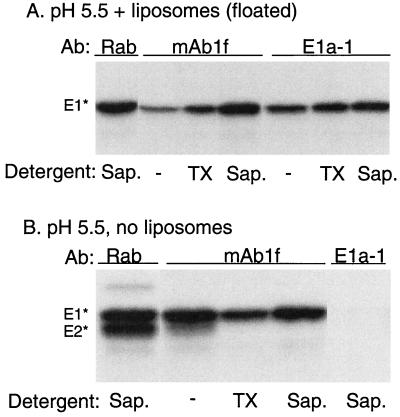
Semliki Forest virus (SFV) is an enveloped alphavirus whose membrane fusion is triggered by low pH and promoted by cholesterol and sphingolipid in the target membrane. Fusion is mediated by E1, a viral membrane protein containing the putative fusion peptide. Virus mutant studies indicate that SFV's cholesterol dependence is controlled by regions of E1 outside of the fusion peptide. Both E1 and E1*, a soluble ectodomain form of E1, interact with membranes in a reaction dependent on low pH, cholesterol, and sphingolipid and form highly stable homotrimers. Here we have used detergent extraction and gradient floatation experiments to demonstrate that E1* associated selectively with detergent-resistant membrane domains (DRMs or rafts). In contrast, reconstituted full-length E1 protein or influenza virus fusion peptide was not associated with DRMs. Methyl beta-cyclodextrin quantitatively extracted both cholesterol and E1* from membranes in the absence of detergent, suggesting a strong association of E1* with sterol. Monoclonal antibody studies demonstrated that raft association was mediated by the proposed E1 fusion peptide. Thus, although other regions of E1 are implicated in the control of virus cholesterol dependence, once the SFV fusion peptide inserts in the target membrane it has a high affinity for membrane domains enriched in cholesterol and sphingolipid.
Figures
References
-
- Ahmed, S. N., D. A. Brown, and E. London. 1997. On the origin of sphingolipid/cholesterol-rich detergent-insoluble cell membranes: physiological concentrations of cholesterol and sphingolipid induce formation of a detergent-insoluble, liquid-ordered lipid phase in model membranes. Biochemistry 36:10944-10953. - PubMed
-
- Anderson, R. G. W. 1998. The caveolae membrane system. Annu. Rev. Biochem. 67:199-225. - PubMed
-
- Brown, D. A., and E. London. 1998. Functions of lipid rafts in biological membranes. Annu. Rev. Cell Dev. Biol. 14:111-136. - PubMed
-
- Brown, D. A., and E. London. 2000. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J. Biol. Chem. 275:17221-17224. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
