

doi: 10.1128/jvi.76.7.3579-3584.2002.
Inhibition of in vitro leukocyte proliferation by morbilliviruses
Affiliations
- PMID: 11884584
- PMCID: PMC136012
- DOI: 10.1128/jvi.76.7.3579-3584.2002
Item in Clipboard
Inhibition of in vitro leukocyte proliferation by morbilliviruses
J Virol.
2002 Apr.
Abstract
Immune suppression associated with morbillivirus infections may influence the mortality rate by allowing secondary bacterial infections that are lethal to the host to flourish. Using an in vitro proliferation assay, we have shown that all members of the genus Morbillivirus inhibit the proliferation of a human B-lymphoblast cell line (BJAB). Proliferation of freshly isolated, stimulated bovine and caprine peripheral blood lymphocytes is also inhibited by UV-inactivated rinderpest (RPV) and peste-des-petits ruminants viruses. As for measles virus, coexpression of both the fusion and the hemagglutinin proteins of RPV is necessary and sufficient to induce immune suppression in vitro.
Figures
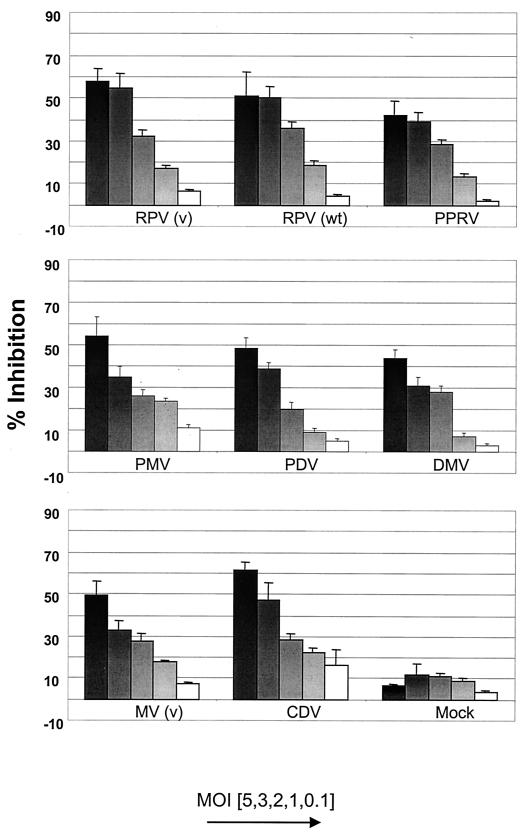
Effect of morbilliviruses on BJAB cell proliferation. Various amounts of UV-inactivated morbilliviruses, corresponding to original MOIs from 0.1 to 5, were mixed with BJAB cells (RC) for 72 h. Proliferation of the RC was determined by the MTT assay, and inhibition induced by the inactive virus was calculated according to the formula [(optical density at 550 nm of control cells − optical density at 550 nm of inactive virus and cells)/optical density at 550 nm of control cells] × 100.
Effect of other viruses and soluble factors on BJAB cell proliferation. (A) Virus preparations, excluding FMDV empty capsids, were UV inactivated and mixed at an MOI of 5 with BJAB cells for 72 h. The empty capsids of FMDV A10 were prepared in a suspension of storage buffer (0.75% ammonium persulfate, 50 mM phosphate, 20 mM Tris [pH 7.5]) at a concentration of 0.73 mg/ml. (B) A PC population was generated by infecting BJAB cells with MV (v) or RPV (v) at an MOI of 3 for 48 h. PC and uninfected BJAB cells (RC) were either mixed (−) or separated by a porous membrane (+) and incubated for 72 h. In each case, proliferation of the RC was determined as described for Fig. 1.
Effect of RPV and PPRV on host PBL proliferation. Freshly isolated caprine and bovine PBL were stimulated with phytohemagglutinin and cultured with UV-inactivated PPRV and RPV vaccine strains (v) or wild-type (wt) RPV and vaccine RPV strains, respectively, at an MOI from 0.1 to 5 for 72 h. In each case, proliferation of the RC was determined with the MTT assay and inhibition was calculated as described for Fig. 1.
Effect of capripoxvirus recombinants expressing RPV H and F proteins on BJAB cell proliferation. PC were generated by infecting BJAB cells at an MOI of 1 with RPV protein expressing recombinant capripoxviruses for 48 h. Cells were either infected singly or with both recombinant viruses. PC were mixed with uninfected BJAB cells (RC) for 72 h; then RC proliferation determined as described for Fig. 1.
Effect of adenovirus recombinants expressing RPV proteins on BJAB cell proliferation. (A) The levels of expression of RPV H and F proteins were determined by fluorescence-activated cell sorting analysis using a rabbit anti-RPV hyperimmune serum. For staining, cells were first washed three times with phosphate-buffered saline containing 1% fetal calf serum and 0.1% sodium azide (PBA). They were then incubated with the antibody for 45 min at 4°C, washed twice with PBA, and then incubated with fluorescein isothiocyanate-conjugated goat anti-rabbit immunoglobulin G (Molecular Probes) for 45 min at 4°C. The cells were then washed and resuspended in PBA for FACScan (Becton Dickinson) analysis with Cell Quest software. (B) PC were generated by infecting BJAB cells at an MOI of 500 with RPV protein expressing recombinant adenovirus vectors for 48 h. The recombinant proteins were either expressed alone or coexpressed in pairs; e.g., where H and F were coexpressed in the same cell, the designation is H/F. Where a heterologous PC population was generated by mixing equal numbers of cells expressing only one protein, the designation is H+F. PC were mixed with uninfected BJAB cells (RC) for 72 h; then the RC proliferation was determined as described for Fig. 1.
References
-
- Barrett, T. 2001. Morbilliviruses: dangers old and new, p. 155-178. In G. L. Smith, J. W. McCauley, and D. J. Rowlands (ed.), New challenges to health: the threat of virus infection. Society for General Microbiology, Symposium 60. Cambridge University Press, Cambridge, United Kingdom.
-
- Barrett, T., I. K. G. Visser, L. Mamaev, L. Goatley, M. F. van Bressem, and A. D. M. E. Osterhaus. 1993. Dolphin and porpoise morbilliviruses are genetically distinct from phocine distemper virus. Virology 193:1010-1012. - PubMed
-
- Cosby, S. L., S. McQuaid, N. Duffy, C. Lyons, B. K. Rima, G. M. Allen, S. J. McCullough, S. Kennedy, J. A. Smyth, F. McNeilly, C. Craig, and C. Örvell. 1988. Characterisation of a seal morbillivirus. Nature 336:115-116.
-
- Domingo, M., M. Vilafranca, J. Visa, N. Prats, A. Trudgett, and I. Visser. 1995. Evidence for chronic morbillivirus infection in the Mediterranean striped dolphin (Stenella coeruleoalba). Vet. Microbiol. 44:229-239. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
