

hnRNP A1 nucleocytoplasmic shuttling activity is required for normal myelopoiesis and BCR/ABL leukemogenesis
- PMID: 11884611
- PMCID: PMC133663
- DOI: 10.1128/MCB.22.7.2255-2266.2002
hnRNP A1 nucleocytoplasmic shuttling activity is required for normal myelopoiesis and BCR/ABL leukemogenesis
Abstract
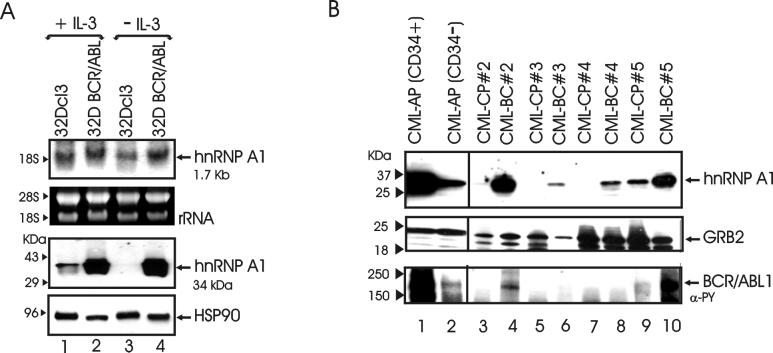
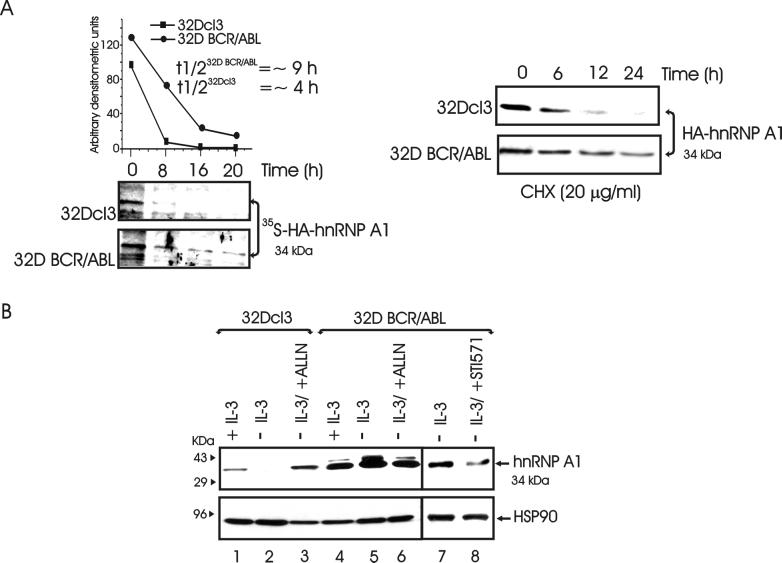
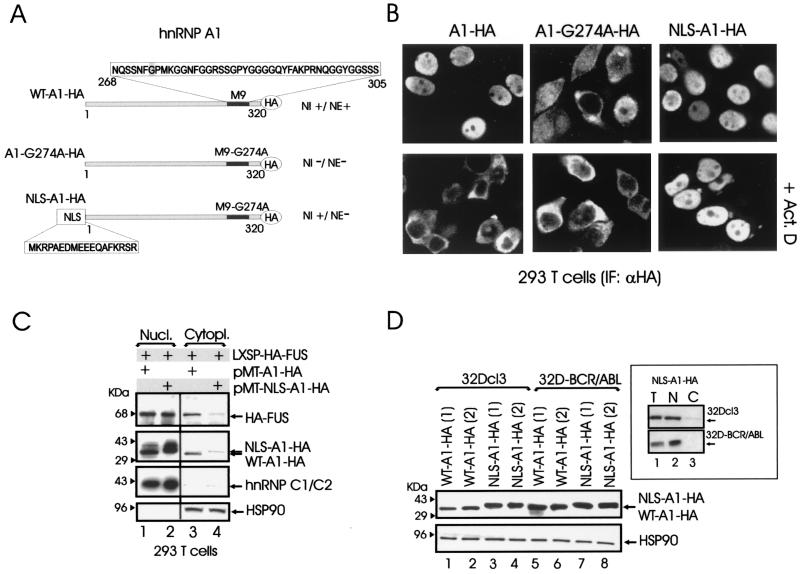
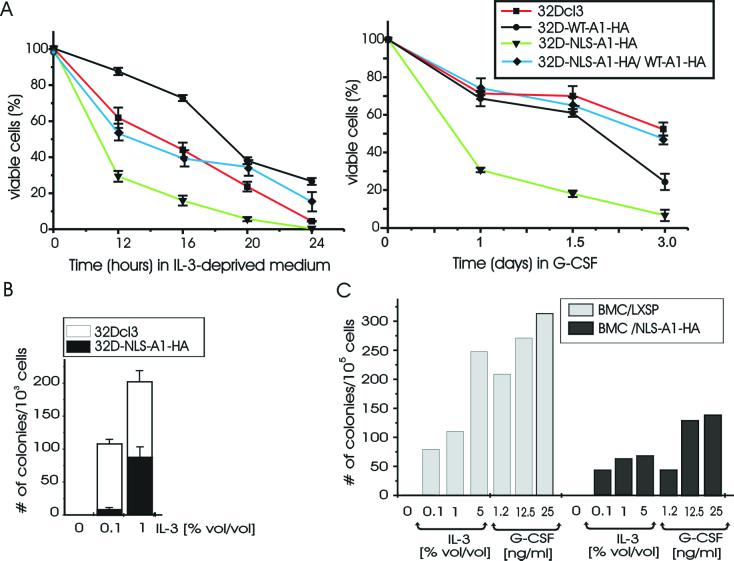
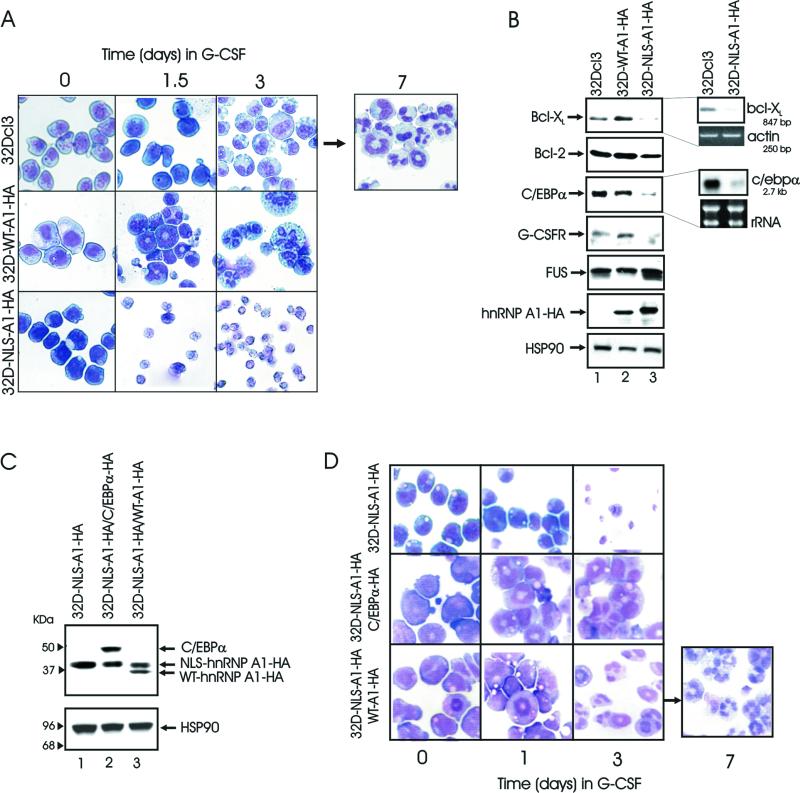
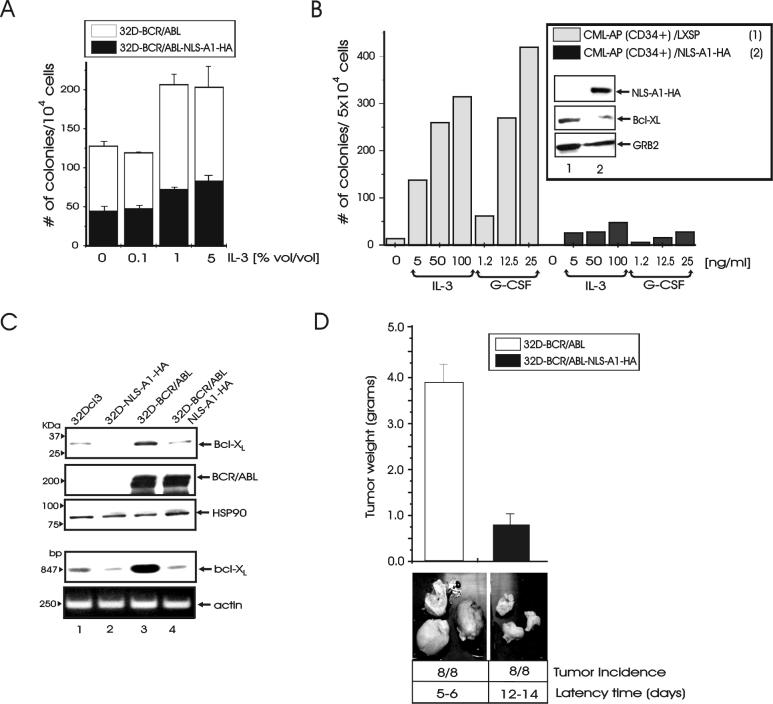
hnRNP A1 is a nucleocytoplasmic shuttling heterogeneous nuclear ribonucleoprotein that accompanies eukaryotic mRNAs from the active site of transcription to that of translation. Although the importance of hnRNP A1 as a regulator of nuclear pre-mRNA and mRNA processing and export is well established, it is unknown whether this is relevant for the control of proliferation, survival, and differentiation of normal and transformed cells. We show here that hnRNP A1 levels are increased in myeloid progenitor cells expressing the p210(BCR/ABL) oncoprotein, in mononuclear cells from chronic myelogenous leukemia (CML) blast crisis patients, and during disease progression. In addition, in myeloid progenitor 32Dcl3 cells, BCR/ABL stabilizes hnRNP A1 by preventing its ubiquitin/proteasome-dependent degradation. To assess the potential role of hnRNP A1 nucleocytoplasmic shuttling activity in normal and leukemic myelopoiesis, a mutant defective in nuclear export was ectopically expressed in parental and BCR/ABL-transformed myeloid precursor 32Dcl3 cells, in normal murine marrow cells, and in mononuclear cells from a CML patient in accelerated phase. In normal cells, expression of this mutant enhanced the susceptibility to apoptosis induced by interleukin-3 deprivation, suppressed granulocytic differentiation, and induced massive cell death of granulocyte colony-stimulating factor-treated cultures. In BCR/ABL-transformed cells, its expression was associated with suppression of colony formation and reduced tumorigenic potential in vivo. Moreover, interference with hnRNP A1 shuttling activity resulted in downmodulation of C/EBPalpha, the major regulator of granulocytic differentiation, and Bcl-X(L), an important survival factor for hematopoietic cells. Together, these results suggest that the shuttling activity of hnRNP A1 is important for the nucleocytoplasmic trafficking of mRNAs that encode proteins influencing the phenotype of normal and BCR/ABL-transformed myeloid progenitors.
Figures
Similar articles
-
High levels of the BCR/ABL oncoprotein are required for the MAPK-hnRNP-E2 dependent suppression of C/EBPalpha-driven myeloid differentiation.Blood. 2007 Aug 1;110(3):994-1003. doi: 10.1182/blood-2007-03-078303. Epub 2007 May 2. Blood. 2007. PMID: 17475908 Free PMC article.
-
BCR-ABL prevents c-jun-mediated and proteasome-dependent FUS (TLS) proteolysis through a protein kinase CbetaII-dependent pathway.Mol Cell Biol. 2000 Aug;20(16):6159-69. doi: 10.1128/MCB.20.16.6159-6169.2000. Mol Cell Biol. 2000. PMID: 10913197 Free PMC article.
-
Identification of novel posttranscriptional targets of the BCR/ABL oncoprotein by ribonomics: requirement of E2F3 for BCR/ABL leukemogenesis.Blood. 2008 Jan 15;111(2):816-28. doi: 10.1182/blood-2007-05-090472. Epub 2007 Oct 9. Blood. 2008. PMID: 17925491 Free PMC article.
-
Post-transcriptional mechanisms in BCR/ABL leukemogenesis: role of shuttling RNA-binding proteins.Oncogene. 2002 Dec 9;21(56):8577-83. doi: 10.1038/sj.onc.1206085. Oncogene. 2002. PMID: 12476304 Review.
-
From mRNA metabolism to cancer therapy: chronic myelogenous leukemia shows the way.Clin Cancer Res. 2007 Mar 15;13(6):1638-42. doi: 10.1158/1078-0432.CCR-06-2320. Clin Cancer Res. 2007. PMID: 17363515 Review.
Cited by
-
A comprehensive understanding of hnRNP A1 role in cancer: new perspectives on binding with noncoding RNA.Cancer Gene Ther. 2023 Mar;30(3):394-403. doi: 10.1038/s41417-022-00571-1. Epub 2022 Dec 2. Cancer Gene Ther. 2023. PMID: 36460805 Review.
-
Human immunodeficiency virus type 1 (HIV-1) induces the cytoplasmic retention of heterogeneous nuclear ribonucleoprotein A1 by disrupting nuclear import: implications for HIV-1 gene expression.J Biol Chem. 2009 Nov 6;284(45):31350-62. doi: 10.1074/jbc.M109.048736. Epub 2009 Sep 8. J Biol Chem. 2009. PMID: 19737937 Free PMC article.
-
hnRNPA1 couples nuclear export and translation of specific mRNAs downstream of FGF-2/S6K2 signalling.Nucleic Acids Res. 2014 Nov 10;42(20):12483-97. doi: 10.1093/nar/gku953. Epub 2014 Oct 16. Nucleic Acids Res. 2014. PMID: 25324306 Free PMC article.
-
High levels of the BCR/ABL oncoprotein are required for the MAPK-hnRNP-E2 dependent suppression of C/EBPalpha-driven myeloid differentiation.Blood. 2007 Aug 1;110(3):994-1003. doi: 10.1182/blood-2007-03-078303. Epub 2007 May 2. Blood. 2007. PMID: 17475908 Free PMC article.
-
MNK1 expression increases during cellular senescence and modulates the subcellular localization of hnRNP A1.Exp Cell Res. 2012 Mar 10;318(5):500-8. doi: 10.1016/j.yexcr.2011.12.015. Epub 2011 Dec 27. Exp Cell Res. 2012. PMID: 22227431 Free PMC article.
References
-
- Amarante-Mendes, G. P., A. J. McGahon, W. K. Nishioka, D. E. Afar, O. N. Witte, and D. R. Green. 1998. Bcl-2-independent Bcr-Abl-mediated resistance to apoptosis: protection is correlated with up regulation of Bcl-xL. Oncogene 16:1383-1390. - PubMed
-
- Bellon, T., D. Perrotti, and B. Calabretta. 1997. Granulocytic differentiation of normal hematopoietic precursor cells induced by transcription factor PU.1 correlates with negative regulation of the c-myb promoter. Blood 90:1828-1839. - PubMed
-
- Biamonti, G., M. T. Bassi, L. Cartegni, F. Mechta, M. Buvoli, F. Cobianchi, and S. Riva. 1993. Human hnRNP protein A1 gene expression. Structural and functional characterization of the promoter. J. Mol. Biol. 230:77-89. - PubMed
-
- Biamonti, G., M. Buvoli, M. T. Bassi, C. Morandi, F. Cobianchi, and S. Riva. 1989. Isolation of an active gene encoding human hnRNP protein A1. Evidence for alternative splicing. J. Mol. Biol. 207:491-503. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous