

Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar body formation, and photomorphogenesis
- PMID: 11884677
- PMCID: PMC152915
- DOI: 10.1105/tpc.010302
Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar body formation, and photomorphogenesis
Abstract
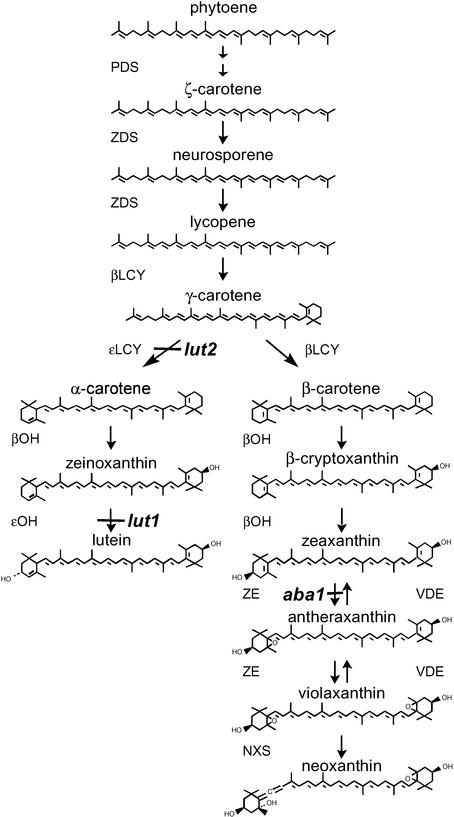
Carotenoids are essential photoprotective and antioxidant pigments synthesized by all photosynthetic organisms. Most carotenoid biosynthetic enzymes were thought to have evolved independently in bacteria and plants. For example, in bacteria, a single enzyme (CrtI) catalyzes the four desaturations leading from the colorless compound phytoene to the red compound lycopene, whereas plants require two desaturases (phytoene and zeta-carotene desaturases) that are unrelated to the bacterial enzyme. We have demonstrated that carotenoid desaturation in plants requires a third distinct enzyme activity, the carotenoid isomerase (CRTISO), which, unlike phytoene and zeta-carotene desaturases, apparently arose from a progenitor bacterial desaturase. The Arabidopsis CRTISO locus was identified by the partial inhibition of lutein synthesis in light-grown tissue and the accumulation of poly-cis-carotene precursors in dark-grown tissue of crtISO mutants. After positional cloning, enzymatic analysis of CRTISO expressed in Escherichia coli confirmed that the enzyme catalyzes the isomerization of poly-cis-carotenoids to all-trans-carotenoids. Etioplasts of dark-grown crtISO mutants accumulate acyclic poly-cis-carotenoids in place of cyclic all-trans-xanthophylls and also lack prolamellar bodies (PLBs), the lattice of tubular membranes that defines an etioplast. This demonstrates a requirement for carotenoid biosynthesis to form the PLB. The absence of PLBs in crtISO mutants demonstrates a function for this unique structure and carotenoids in facilitating chloroplast development during the first critical days of seedling germination and photomorphogenesis.
Figures








Comment in
-
Tangerine dreams: cloning of carotenoid isomerase from Arabidopsis and tomato.Plant Cell. 2002 Feb;14(2):289-92. doi: 10.1105/tpc.140210. Plant Cell. 2002. PMID: 11884674 Free PMC article. No abstract available.
Similar articles
-
Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of beta-carotene and xanthophylls in plants.Plant Cell. 2002 Feb;14(2):333-42. doi: 10.1105/tpc.010303. Plant Cell. 2002. PMID: 11884678 Free PMC article.
-
Dissection of tomato lycopene biosynthesis through virus-induced gene silencing.Plant Physiol. 2013 Oct;163(2):986-98. doi: 10.1104/pp.113.224733. Epub 2013 Sep 6. Plant Physiol. 2013. PMID: 24014574 Free PMC article.
-
Two Arabidopsis thaliana carotene desaturases, phytoene desaturase and zeta-carotene desaturase, expressed in Escherichia coli, catalyze a poly-cis pathway to yield pro-lycopene.Eur J Biochem. 1999 Jan;259(1-2):396-403. doi: 10.1046/j.1432-1327.1999.00051.x. Eur J Biochem. 1999. PMID: 9914519
-
Carotenoids, versatile components of oxygenic photosynthesis.Prog Lipid Res. 2013 Oct;52(4):539-61. doi: 10.1016/j.plipres.2013.07.001. Epub 2013 Jul 26. Prog Lipid Res. 2013. PMID: 23896007 Review.
-
Evolution of carotene desaturation: the complication of a simple pathway.Arch Biochem Biophys. 2009 Mar 15;483(2):169-74. doi: 10.1016/j.abb.2008.10.004. Epub 2008 Oct 12. Arch Biochem Biophys. 2009. PMID: 18948076 Review.
Cited by
-
Application of 5-aminolevulinic acid promotes ripening and accumulation of primary and secondary metabolites in postharvest tomato fruit.Front Nutr. 2022 Nov 10;9:1036843. doi: 10.3389/fnut.2022.1036843. eCollection 2022. Front Nutr. 2022. PMID: 36438749 Free PMC article.
-
Analysis of carotenoid accumulation and expression of carotenoid biosynthesis genes in different organs of Chinese cabbage (Brassica rapa subsp. pekinensis).EXCLI J. 2012 Aug 22;11:508-16. eCollection 2012. EXCLI J. 2012. PMID: 27540344 Free PMC article.
-
Accumulation of carotenoids and expression of carotenoid biosynthetic genes during maturation in citrus fruit.Plant Physiol. 2004 Feb;134(2):824-37. doi: 10.1104/pp.103.031104. Epub 2004 Jan 22. Plant Physiol. 2004. PMID: 14739348 Free PMC article.
-
A foliar pigment-based bioassay for interrogating chloroplast signalling revealed that carotenoid isomerisation regulates chlorophyll abundance.Plant Methods. 2022 Feb 17;18(1):18. doi: 10.1186/s13007-022-00847-5. Plant Methods. 2022. PMID: 35177117 Free PMC article.
-
Carotenoid modifying enzymes in metazoans.Methods Enzymol. 2022;674:405-445. doi: 10.1016/bs.mie.2022.05.005. Epub 2022 Jun 22. Methods Enzymol. 2022. PMID: 36008015 Free PMC article.
References
-
- Axelsson, L., Dahlin, C., and Ryberg, H. (1982). The function of carotenoids during chloroplast development. V. Correlation be-tween carotenoid content, ultrastructure and chlorophyll b to chlorophyll a ratio. Physiol. Plant. 55, 111–116.
-
- Bartley, G.E., Scolnik, P.A., and Beyer, P. (1999). Two Arabidopsis thaliana carotene desaturases, phytoene desaturase and zeta-carotene desaturase, expressed in Escherichia coli, catalyze a poly-cis pathway to yield pro-lycopene. Eur. J. Biochem. 259, 396–403. - PubMed
-
- Bell, C.J., and Ecker, J.R. (1994). Assignment of 30 microsatellite loci to the linkage map of Arabidopsis. Genomics 19, 137–144. - PubMed
-
- Beyer, P., Mayer, M., and Kleinig, H. (1989). Molecular oxygen and the state of geometric isomerism of intermediates are essential in the carotene desaturation and cyclization reactions in daffodil chromoplasts. Eur. J. Biochem. 184, 141–150. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
