

Orientation sensitivity of ganglion cells in primate retina
- PMID: 11888534
- PMCID: PMC6880403
- DOI: 10.1016/s0042-6989(01)00312-1
Orientation sensitivity of ganglion cells in primate retina
Abstract
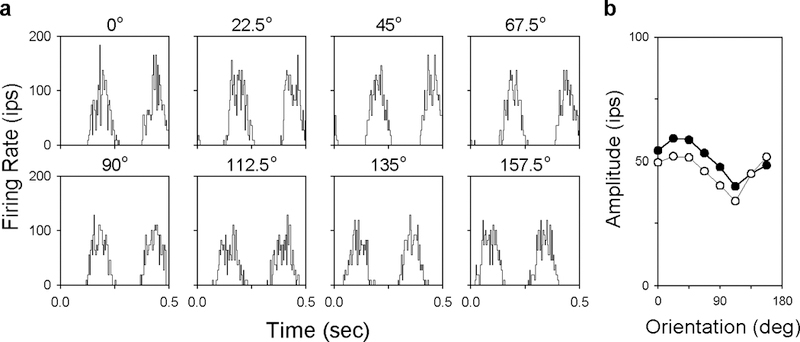
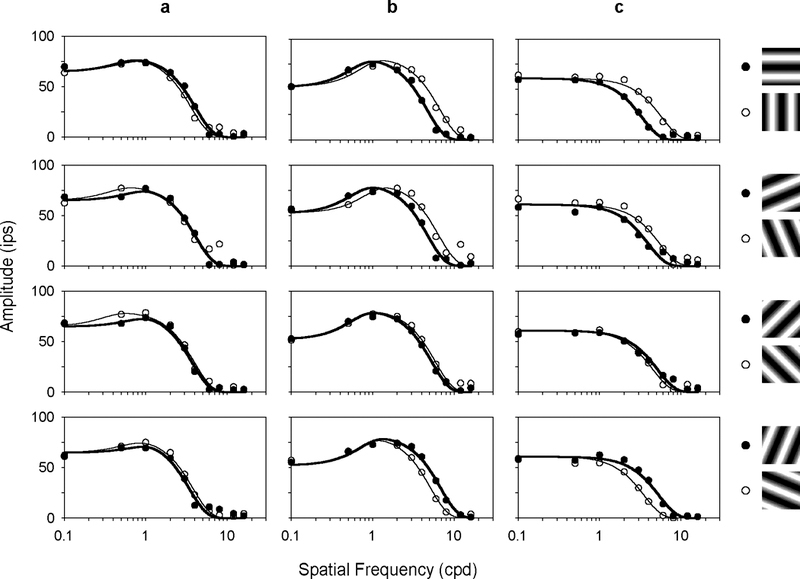
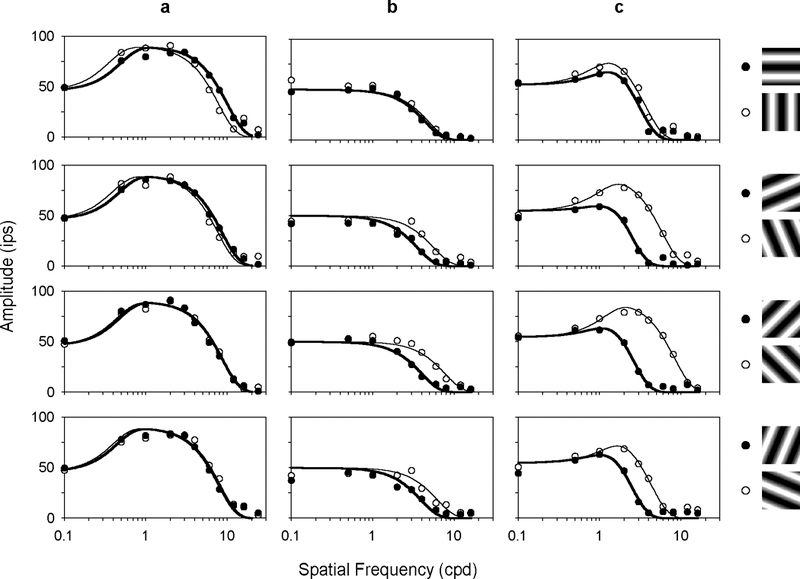
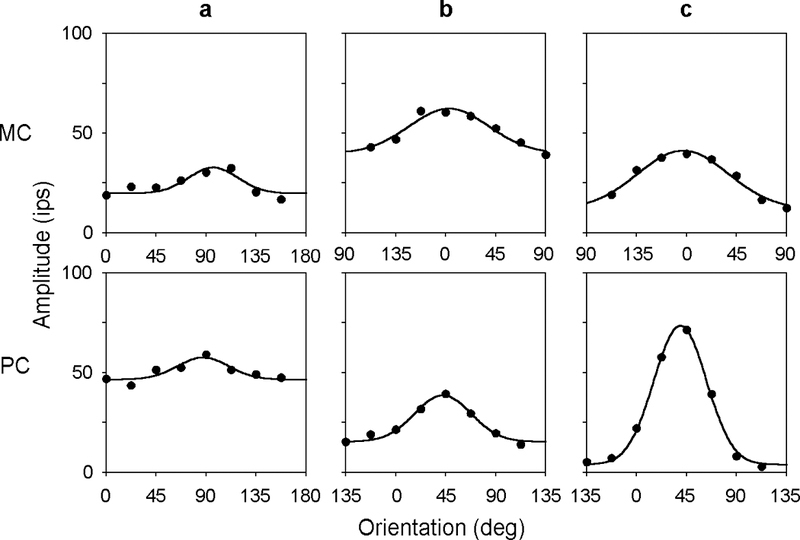
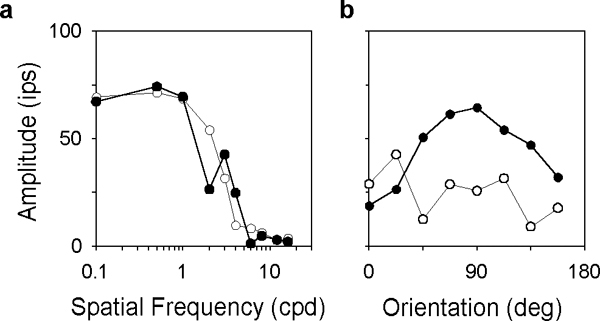
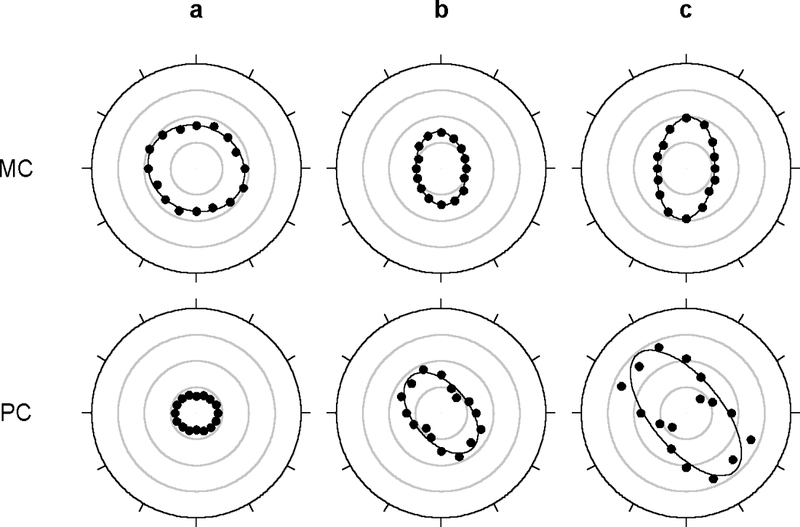
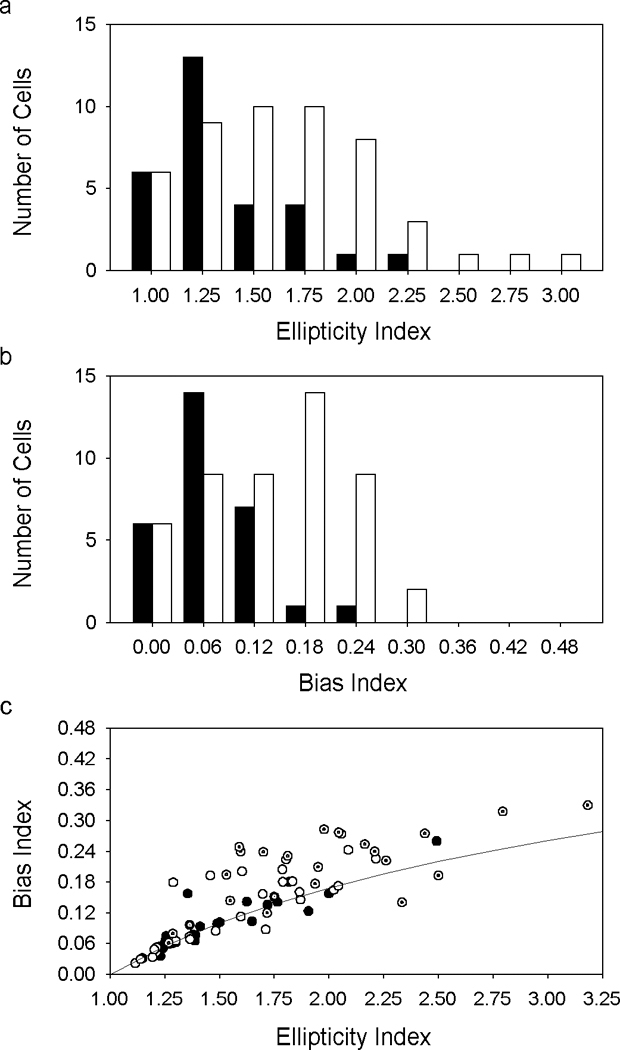
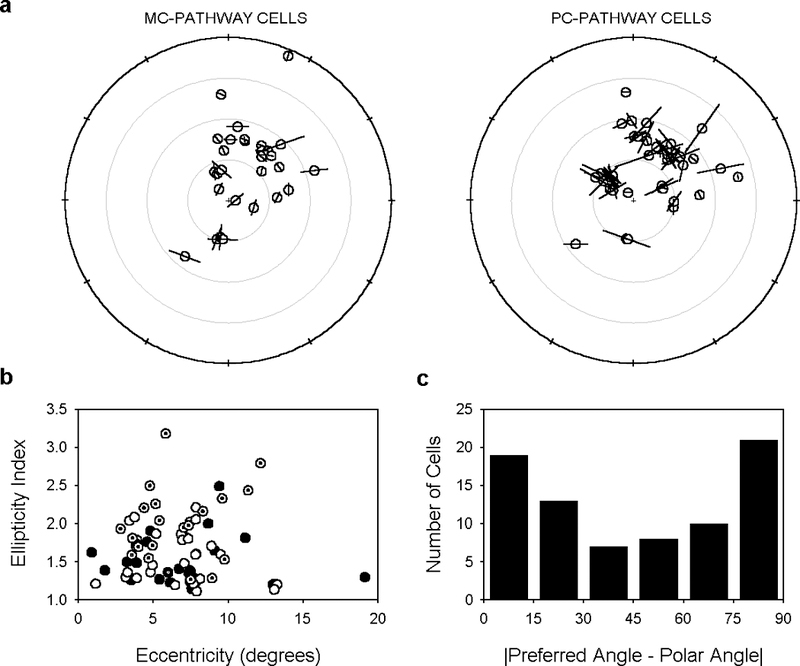
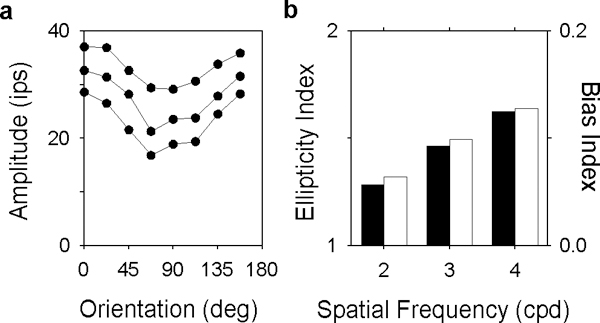
The two-dimensional shape of the receptive field center of macaque retinal ganglion cells was determined by measuring responses to drifting sinusoidal gratings of different spatial frequency and orientation. The responses of most cells to high spatial frequencies depended on grating orientation, indicating that their centers were not circularly symmetric. In general, center shape was well described by an ellipse. The major axis of the ellipse tended to point towards the fovea or perpendicular to this. Parvocellular pathway cells had greater center ellipticity than magnocellular pathway cells; the median ratio of the major-to-minor axis was 1.72 and 1.38, respectively. Parvocellular pathway cells also had centers that were often bimodal in shape, suggesting that they received patchy cone/bipolar cell input. We conclude that most ganglion cells in primate retina have elongated receptive field centers and thus show orientation sensitivity.
Figures
Similar articles
-
L and M cone contributions to the midget and parasol ganglion cell receptive fields of macaque monkey retina.J Neurosci. 2004 Feb 4;24(5):1079-88. doi: 10.1523/JNEUROSCI.3828-03.2004. J Neurosci. 2004. PMID: 14762126 Free PMC article.
-
The spatial structure of cone-opponent receptive fields in macaque retina.Vision Res. 2018 Oct;151:141-151. doi: 10.1016/j.visres.2017.05.013. Epub 2017 Jul 19. Vision Res. 2018. PMID: 28709923
-
Chromatic organization of ganglion cell receptive fields in the peripheral retina.J Neurosci. 2005 May 4;25(18):4527-39. doi: 10.1523/JNEUROSCI.3921-04.2005. J Neurosci. 2005. PMID: 15872100 Free PMC article.
-
Pathways mediating resolution in the primate retina.Ciba Found Symp. 1990;155:5-14; discussion 14-21. doi: 10.1002/9780470514023.ch2. Ciba Found Symp. 1990. PMID: 2088681 Review.
-
Physiology, morphology and spatial densities of identified ganglion cell types in primate retina.Ciba Found Symp. 1994;184:12-28; discussion 28-34, 63-70. doi: 10.1002/9780470514610.ch2. Ciba Found Symp. 1994. PMID: 7882750 Review.
Cited by
-
Probing Computation in the Primate Visual System at Single-Cone Resolution.Annu Rev Neurosci. 2019 Jul 8;42:169-186. doi: 10.1146/annurev-neuro-070918-050233. Epub 2019 Mar 11. Annu Rev Neurosci. 2019. PMID: 30857477 Free PMC article. Review.
-
High-Fidelity Reproduction of Visual Signals by Electrical Stimulation in the Central Primate Retina.J Neurosci. 2023 Jun 21;43(25):4625-4641. doi: 10.1523/JNEUROSCI.1091-22.2023. Epub 2023 May 15. J Neurosci. 2023. PMID: 37188516 Free PMC article.
-
Orientation-Selective Retinal Circuits in Vertebrates.Front Neural Circuits. 2018 Feb 7;12:11. doi: 10.3389/fncir.2018.00011. eCollection 2018. Front Neural Circuits. 2018. PMID: 29467629 Free PMC article. Review.
-
Specificity of M and L cone inputs to receptive fields in the parvocellular pathway: random wiring with functional bias.J Neurosci. 2006 Oct 25;26(43):11148-61. doi: 10.1523/JNEUROSCI.3237-06.2006. J Neurosci. 2006. PMID: 17065455 Free PMC article.
-
Subtype-dependent postnatal development of direction- and orientation-selective retinal ganglion cells in mice.J Neurophysiol. 2014 Nov 1;112(9):2092-101. doi: 10.1152/jn.00320.2014. Epub 2014 Aug 6. J Neurophysiol. 2014. PMID: 25098962 Free PMC article.
References
-
- Bloomfield SA (1994) Orientation-sensitive amacrine and ganglion cells in the rabbit retina. Journal of Neurophysiology, 71, 1672–1691. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
