

Conserved segments 1A and 2B of the intermediate filament dimer: their atomic structures and role in filament assembly
- PMID: 11889032
- PMCID: PMC125921
- DOI: 10.1093/emboj/21.6.1255
Conserved segments 1A and 2B of the intermediate filament dimer: their atomic structures and role in filament assembly
Abstract
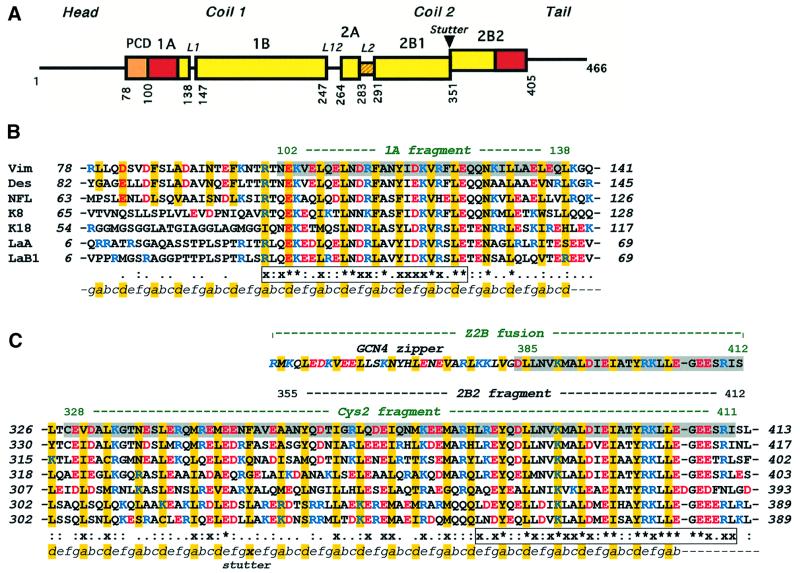
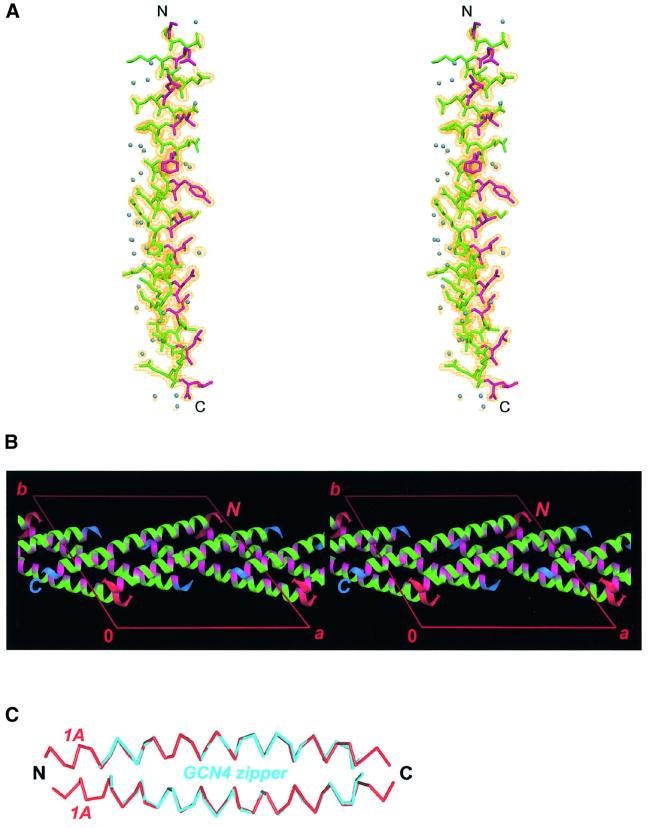
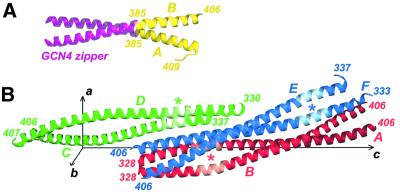
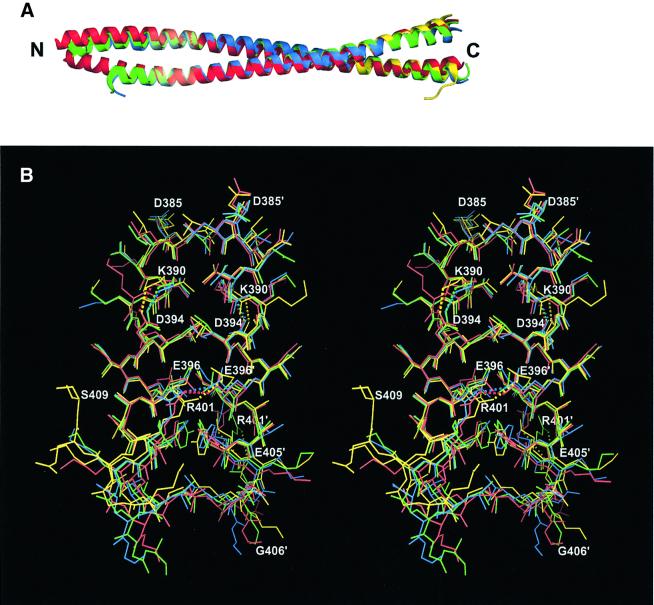
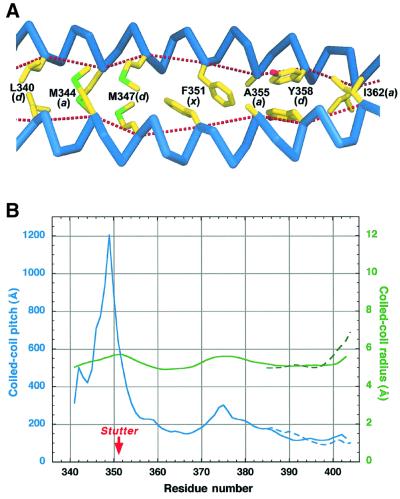
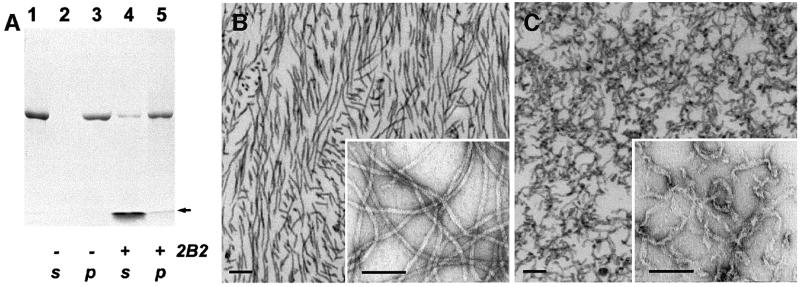
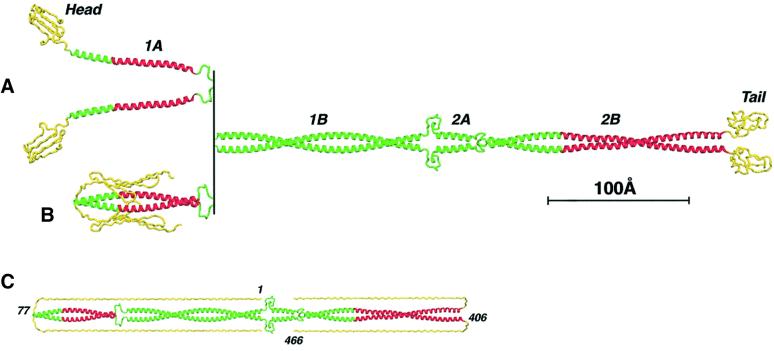
Intermediate filaments (IFs) are key components of the cytoskeleton in higher eukaryotic cells. The elementary IF 'building block' is an elongated coiled-coil dimer consisting of four consecutive alpha-helical segments. The segments 1A and 2B include highly conserved sequences and are critically involved in IF assembly. Based on the crystal structures of three human vimentin fragments at 1.4-2.3 A resolution (PDB entries 1gk4, 1gk6 and 1gk7), we have established the molecular organization of these two segments. The fragment corresponding to segment 1A forms a single, amphipatic alpha-helix, which is compatible with a coiled-coil geometry. While this segment might yield a coiled coil within an isolated dimer, monomeric 1A helices are likely to play a role in specific dimer-dimer interactions during IF assembly. The 2B segment reveals a double-stranded coiled coil, which unwinds near residue Phe351 to accommodate a 'stutter'. A fragment containing the last seven heptads of 2B interferes heavily with IF assembly and also transforms mature vimentin filaments into a new kind of structure. These results provide the first insight into the architecture and functioning of IFs at the atomic level.
Figures
References
-
- Aebi U., Haner,M., Troncoso,J., Eichner,R. and Engel,A. (1988) Unifying principles in intermediate filament structure and assembly. Protoplasma, 145, 73–81.
-
- Brown J.H., Cohen,C. and Parry,D.A. (1996) Heptad breaks in α-helical coiled coils: stutters and stammers. Proteins, 26, 134–145. - PubMed
-
- Brünger A.T. and Nilges,M. (1993) Computational challenges for macromolecular structure determination by X-ray crystallography and solution NMR-spectroscopy. Q. Rev. Biophys., 26, 49–125. - PubMed
-
- Brünger A.T. et al. (1998) Crystallography and NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr. D, 54, 905–921. - PubMed
-
- Burkhard P., Kammerer,R.A., Steinmetz,M.O., Bourenkov,G.P. and Aebi,U. (2000) The coiled-coil trigger site of the rod domain of cortexillin I unveils a distinct network of inter- and intra-helical salt-bridges. Structure, 8, 223–230. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
