

Late endosome motility depends on lipids via the small GTPase Rab7
- PMID: 11889035
- PMCID: PMC125356
- DOI: 10.1093/emboj/21.6.1289
Late endosome motility depends on lipids via the small GTPase Rab7
Abstract
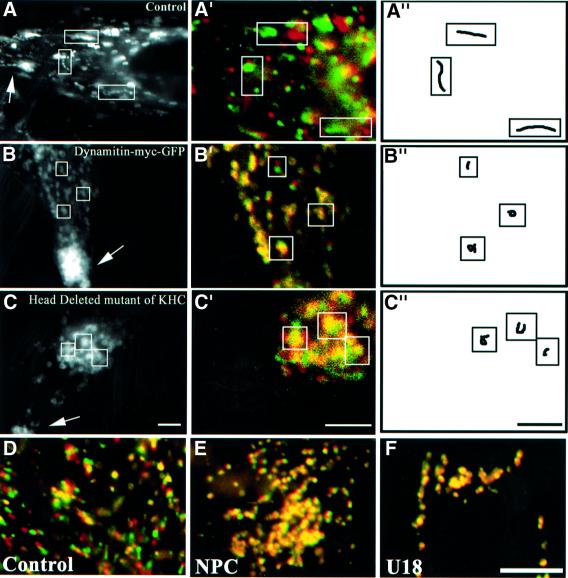
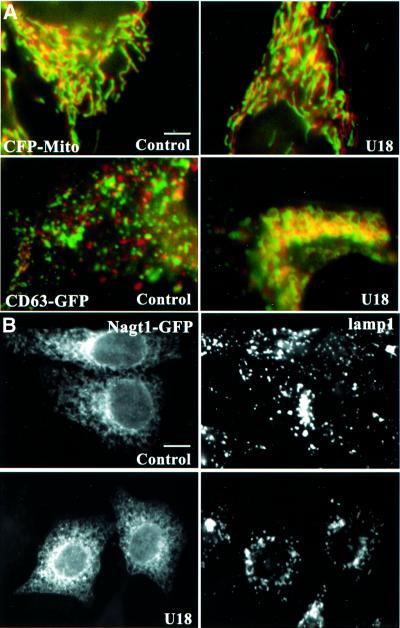
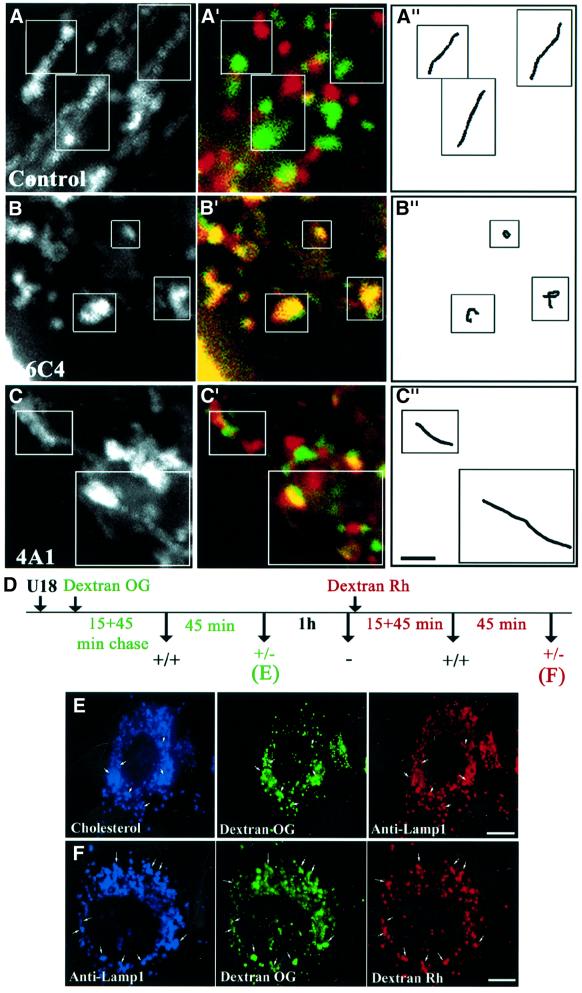
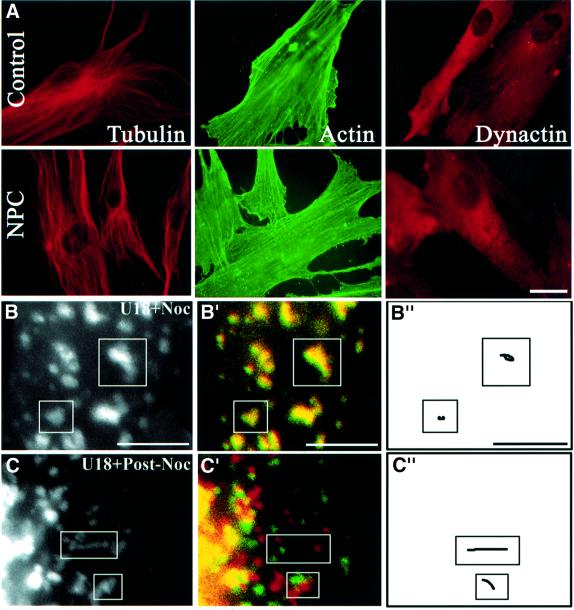
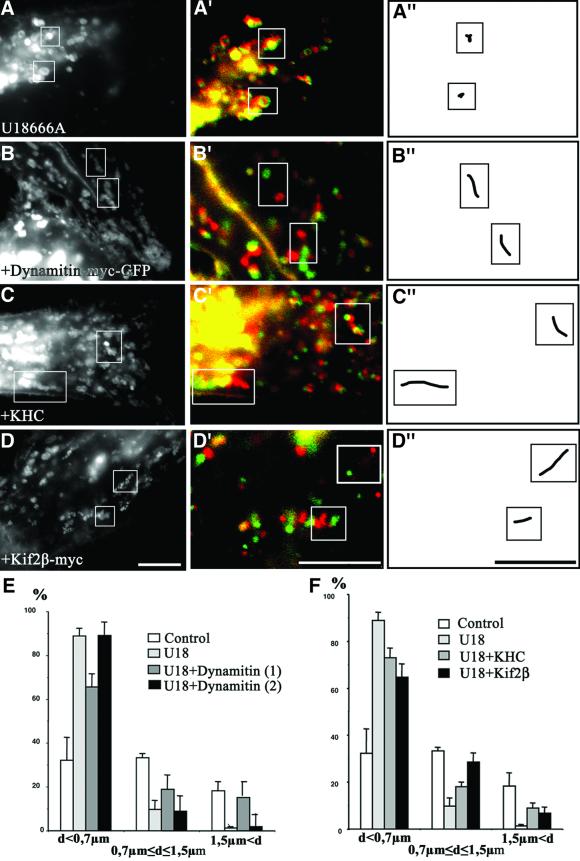
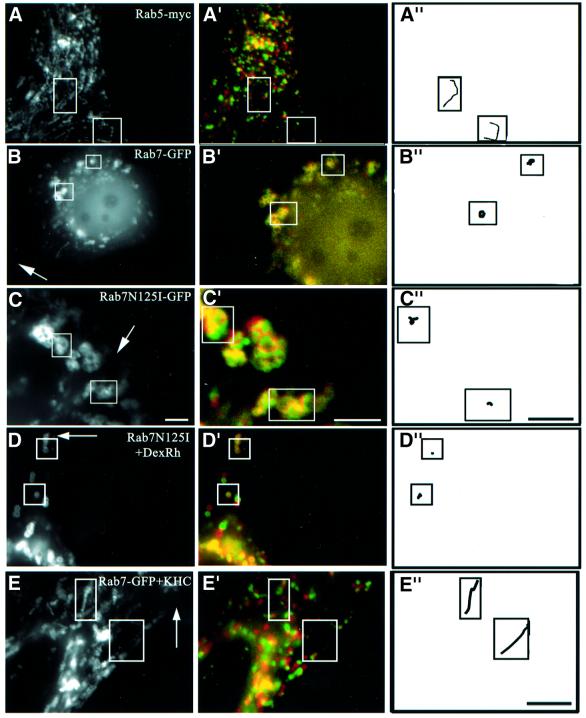
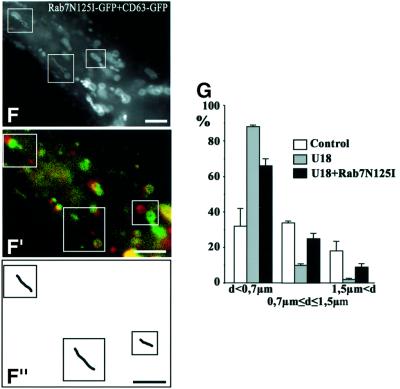
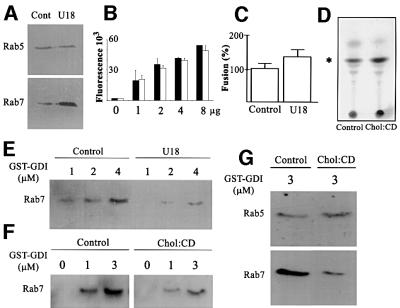
We report that lipids contribute to regulate the bidirectional motility of late endocytic compartments. Late endocytic vesicles loaded with cholesterol lose their dynamic properties, and become essentially immobile, including in cells from Niemann-Pick C patients. These vesicles then retain cytoplasmic dynein activity, but seem to be unable to acquire kinesin activity, eventually leading to paralysis. Our data suggest that this defect depends on the small GTPase Rab7, since the motility of vesicles loaded with cholesterol can be restored by the Rab7 inhibitory mutant N125I. Conversely, wild-type Rab7 overexpression mimics the effects of cholesterol on motility in control cells. Consistently, cholesterol accumulation increases the amounts of membrane-associated Rab7, and inhibits Rab7 membrane extraction by the guanine nucleotide dissociation inhibitor. Our observations thus indicate that cholesterol contributes to regulate the Rab7 cycle, and that Rab7 in turn controls the net movement of late endocytic elements. We conclude that motor functions can be regulated by the membrane lipid composition via the Rab7 cycle.
Figures
Similar articles
-
Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7-RILP-p150 Glued and late endosome positioning.J Cell Biol. 2009 Jun 29;185(7):1209-25. doi: 10.1083/jcb.200811005. J Cell Biol. 2009. PMID: 19564404 Free PMC article.
-
Phagosomes fuse with late endosomes and/or lysosomes by extension of membrane protrusions along microtubules: role of Rab7 and RILP.Mol Cell Biol. 2003 Sep;23(18):6494-506. doi: 10.1128/MCB.23.18.6494-6506.2003. Mol Cell Biol. 2003. PMID: 12944476 Free PMC article.
-
Activation of endosomal dynein motors by stepwise assembly of Rab7-RILP-p150Glued, ORP1L, and the receptor betalll spectrin.J Cell Biol. 2007 Feb 12;176(4):459-71. doi: 10.1083/jcb.200606077. Epub 2007 Feb 5. J Cell Biol. 2007. PMID: 17283181 Free PMC article.
-
Rab7: roles in membrane trafficking and disease.Biosci Rep. 2009 Jun;29(3):193-209. doi: 10.1042/BSR20090032. Biosci Rep. 2009. PMID: 19392663 Review.
-
The Lipids of the Early Endosomes: Making Multimodality Work.Chembiochem. 2017 Jun 19;18(12):1053-1060. doi: 10.1002/cbic.201700046. Epub 2017 May 10. Chembiochem. 2017. PMID: 28374483 Review.
Cited by
-
Hydroxypropyl-beta and -gamma cyclodextrins rescue cholesterol accumulation in Niemann-Pick C1 mutant cell via lysosome-associated membrane protein 1.Cell Death Dis. 2018 Oct 3;9(10):1019. doi: 10.1038/s41419-018-1056-1. Cell Death Dis. 2018. PMID: 30282967 Free PMC article.
-
Linking Late Endosomal Cholesterol with Cancer Progression and Anticancer Drug Resistance.Int J Mol Sci. 2022 Jun 29;23(13):7206. doi: 10.3390/ijms23137206. Int J Mol Sci. 2022. PMID: 35806209 Free PMC article. Review.
-
TMEM135 regulates primary ciliogenesis through modulation of intracellular cholesterol distribution.EMBO Rep. 2020 May 6;21(5):e48901. doi: 10.15252/embr.201948901. Epub 2020 Mar 11. EMBO Rep. 2020. PMID: 32157776 Free PMC article.
-
Endosomal Escape of Antisense Oligonucleotides Internalized by Stabilin Receptors Is Regulated by Rab5C and EEA1 During Endosomal Maturation.Nucleic Acid Ther. 2018 Apr;28(2):86-96. doi: 10.1089/nat.2017.0694. Epub 2018 Feb 13. Nucleic Acid Ther. 2018. PMID: 29437530 Free PMC article.
-
Prion infection impairs lysosomal degradation capacity by interfering with rab7 membrane attachment in neuronal cells.Sci Rep. 2016 Feb 11;6:21658. doi: 10.1038/srep21658. Sci Rep. 2016. PMID: 26865414 Free PMC article.
References
-
- Allan V.J. and Schroer,T.A. (1999) Membrane motors. Curr. Opin. Cell Biol., 11, 476–482. - PubMed
-
- Berrueta L., Tirnauer,J.S., Schuyler,S.C., Pellman,D. and Bierer,B.E. (1999) The APC-associated protein EB1 associates with components of the dynactin complex and cytoplasmic dynein intermediate chain. Curr. Biol., 9, 425–428. - PubMed
-
- Bomsel M., Parton,R., Kuznetsov,S.A., Schroer,T.A. and Gruenberg,J. (1990) Microtubule and motor dependent fusion in vitro between apical and basolateral endocytic vesicles from MDCK cells. Cell, 62, 719–731. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
