

CCR4, a 3'-5' poly(A) RNA and ssDNA exonuclease, is the catalytic component of the cytoplasmic deadenylase
- PMID: 11889047
- PMCID: PMC125924
- DOI: 10.1093/emboj/21.6.1414
CCR4, a 3'-5' poly(A) RNA and ssDNA exonuclease, is the catalytic component of the cytoplasmic deadenylase
Abstract
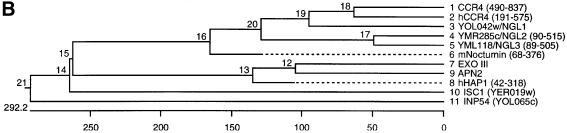
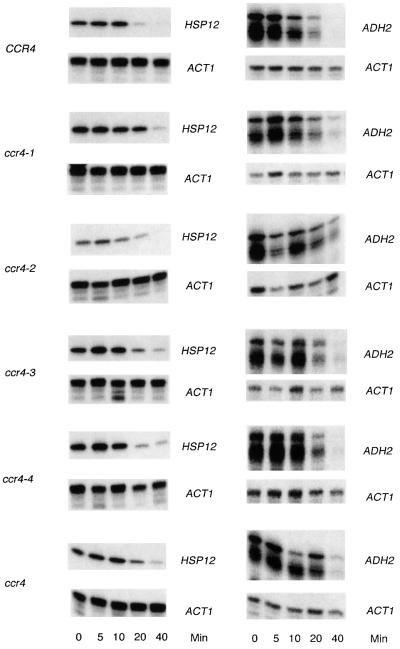
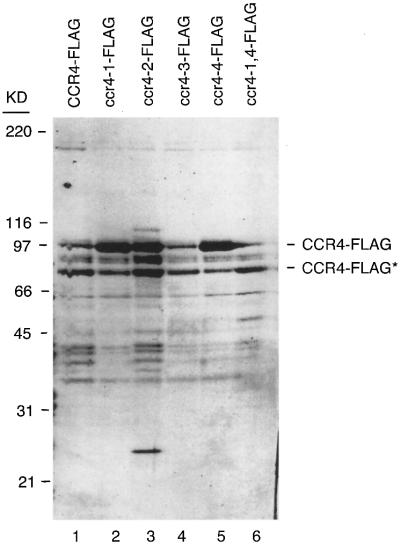
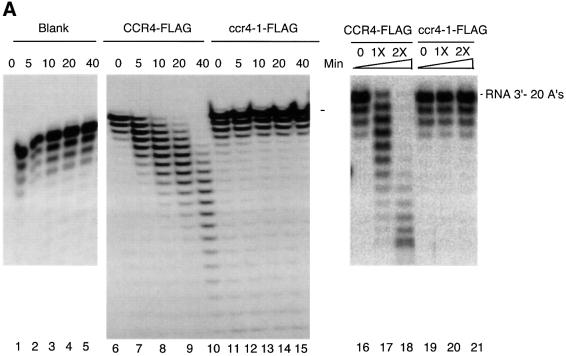
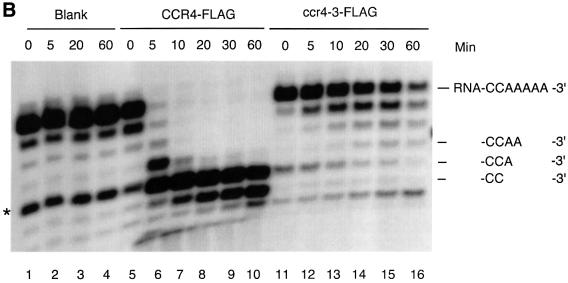
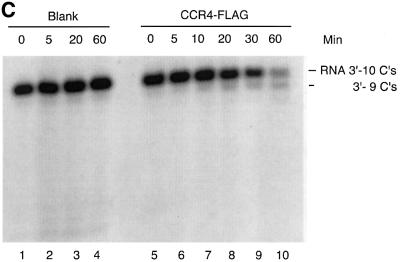
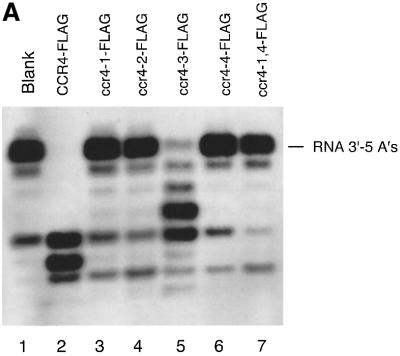
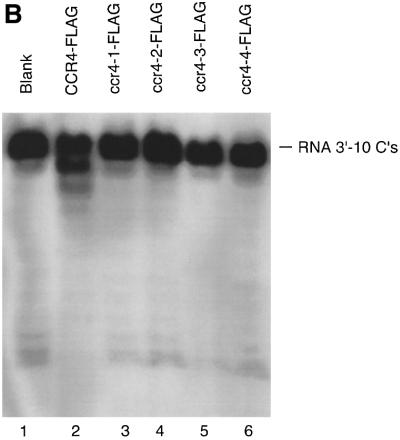
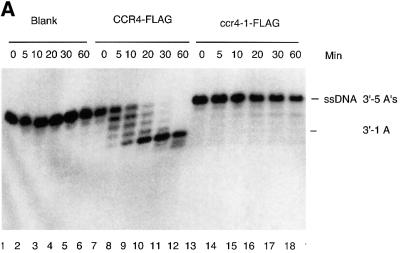
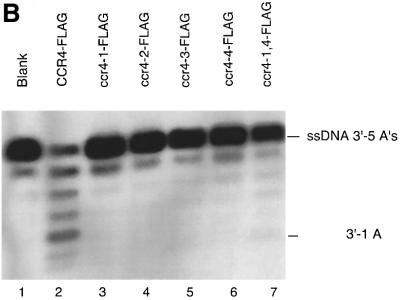
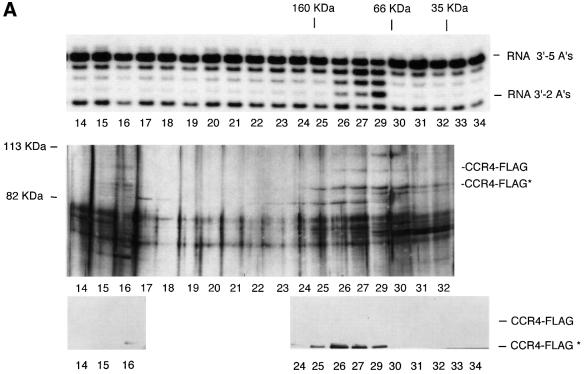
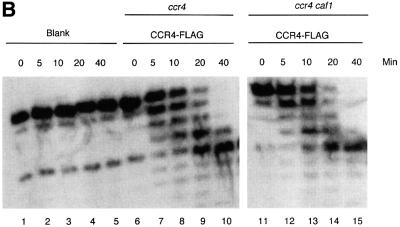
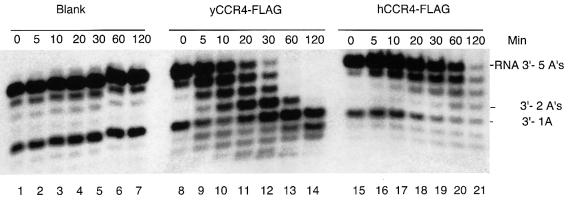
The CCR4-NOT complex from Saccharomyces cerevisiae is a general transcriptional regulatory complex. The proteins of this complex are involved in several aspects of mRNA metabolism, including transcription initiation and elongation and mRNA degradation. The evolutionarily conserved CCR4 protein, which is part of the cytoplasmic deadenylase, contains a C-terminal domain that displays homology to an Mg2+-dependent DNase/phosphatase family of proteins. We have analyzed the putative enzymatic properties of CCR4 and have found that it contains both RNA and single-stranded DNA 3'-5' exonuclease activities. CCR4 displays a preference for RNA and for 3' poly(A) substrates, implicating it as the catalytic component of the cytoplasmic deadenylase. Mutations in the key, conserved catalytic residues in the CCR4 exonuclease domain abolished both its in vitro activities and its in vivo functions. Importantly, CCR4 was active as a monomer and remained active in the absence of CAF1, which links CCR4 to the remainder of the CCR4-NOT complex components. These results establish that CCR4 and most probably other members of a widely distributed CCR4-like family of proteins constitute a novel class of RNA-DNA exonucleases. The various regulatory effects of the CCR4-NOT complex on gene expression may be executed in part through these CCR4 exonuclease activities.
Figures

References
-
- Barzilay G., Mol,C.D., Robson,C.N., Walker,L.J., Cunningham,R.P., Tainer,J.A. and Hickson,I.D. (1995) Identification of critical active-site residues in the multifunctional human DNA repair enzyme HAP1. Nature Struct. Biol., 2, 561–568. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
