

Molecular characterization and mapping of ATOH7, a human atonal homolog with a predicted role in retinal ganglion cell development
- PMID: 11889557
- PMCID: PMC2262845
- DOI: 10.1007/s00335-001-2101-3
Molecular characterization and mapping of ATOH7, a human atonal homolog with a predicted role in retinal ganglion cell development
Abstract
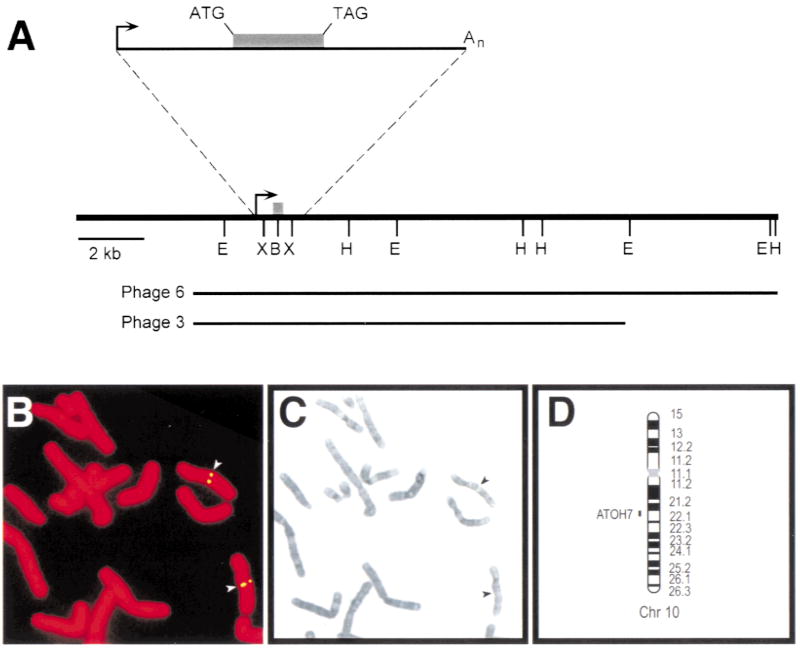
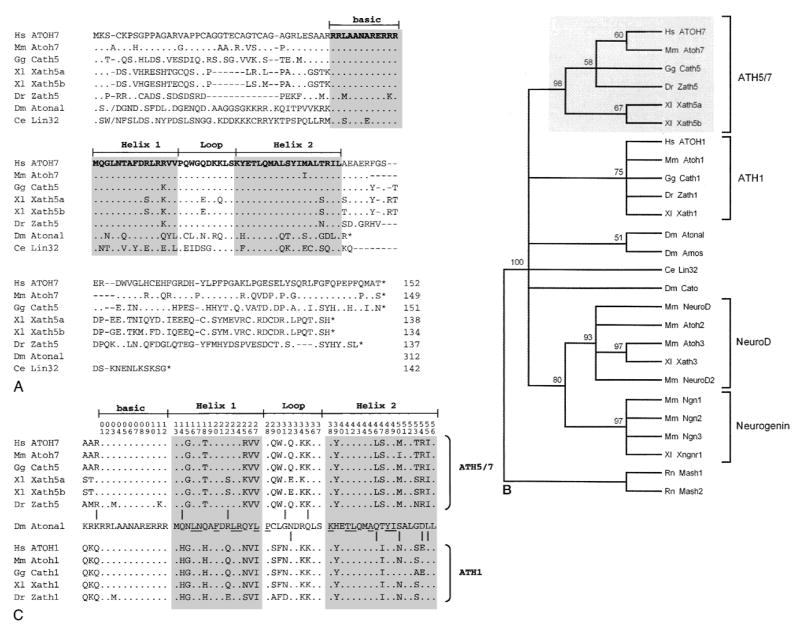
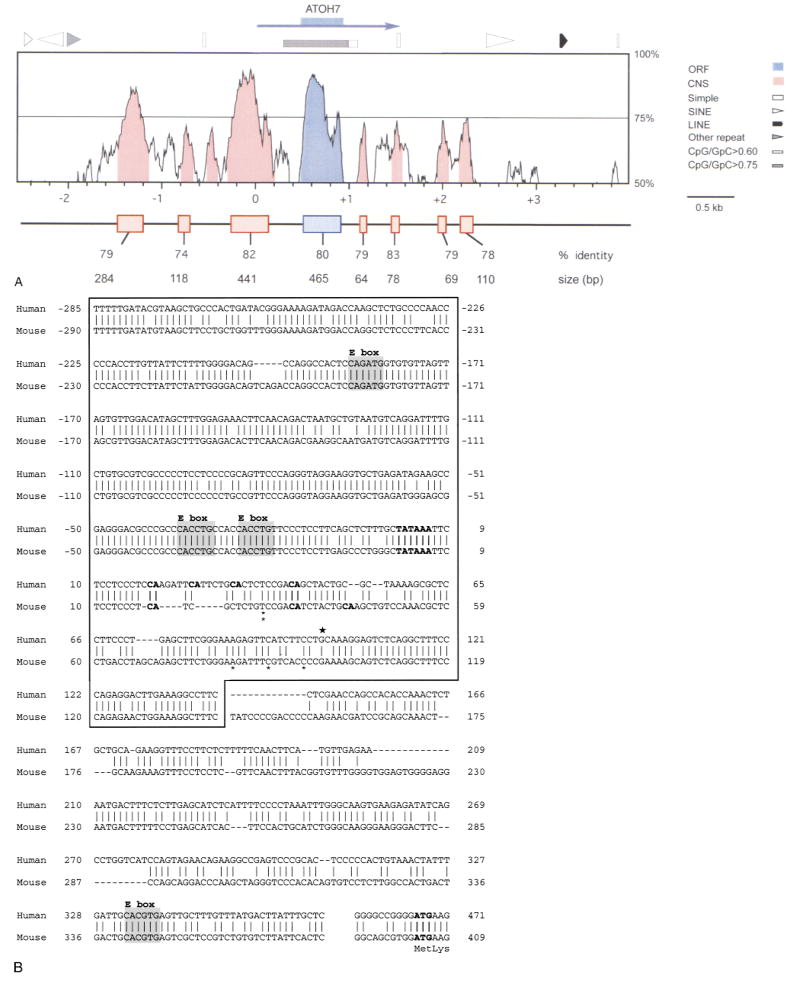
The human ATOH7 gene encodes a basic helix-loop-helix (bHLH) transcription factor that is highly similar to Drosophila Atonal within the conserved bHLH domain. The ATOH7 coding region is contained within a single exon. We mapped ATOH7 to Chromosome (Chr) 10q21.3-22.1, a region syntenic to the segment of mouse Chr 10 where Atoh7 (formerly Math5) is located. The evolutionary relationship between ATOH7 and other atonal homologs was investigated using parsimony analysis. A direct comparison of ATH5/7 and ATH1 protein subgroups to Atonal also revealed a nonrandom distribution of amino acid changes across the bHLH domain, which may be related to their separate visual and proprioceptive sensory functions. Among bHLH genes, ATOH7 is most closely related to Atoh7. This sequence conservation extends significantly beyond the coding region. We define blocks of strong homology in flanking human and mouse genomic DNA, which are likely to include cis regulatory elements. Because targeted deletion of Atoh7 causes optic nerve agenesis in mice, we propose ATOH7 as a candidate for human optic nerve aplasia and related clinical syndromes.
Figures
References
-
- Alagramam KN, Murcia CL, Kwon HY, Pawlowski KS, Wright CG, et al. The mouse Ames waltzer hearing-loss mutant is caused by mutation of Pcdh15, a novel protocadherin gene. Nat Genet. 2001;27:99–102. - PubMed
-
- Atchley WR, Wollenberg KR, Fitch WM, Terhalle W, Dress AW. Correlations among amino acid sites in bHLH protein domains: an information theoretic analysis. Mol Biol Evol. 2000;17:164–178. - PubMed
-
- Bermingham NA, Hassan BA, Price SD, Vollrath MA, Ben-Arie N, et al. Math1: an essential gene for the generation of inner ear hair cells. Science. 1999;284:1837–1841. - PubMed
-
- Bermingham NA, Hassan BA, Wang VY, Fernandez M, Banfi S, et al. Proprioceptor pathway development is dependent on math1. Neuron. 2001;30:411–422. - PubMed
-
- Bolz H, von Brederlow B, Ramirez A, Bryda EC, Kutsche K, et al. Mutation of CDH23, encoding a new member of the cadherin gene family, causes Usher syndrome type 1D. Nat Genet. 2001;27:108–112. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
