

Ferrous ion transport across chloroplast inner envelope membranes
- PMID: 11891257
- PMCID: PMC152214
- DOI: 10.1104/pp.010858
Ferrous ion transport across chloroplast inner envelope membranes
Abstract
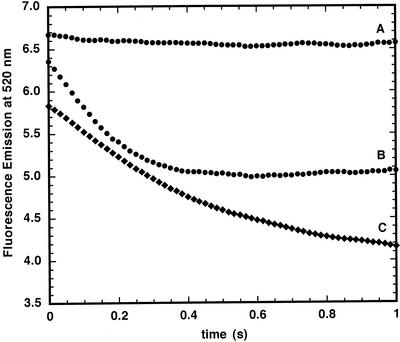
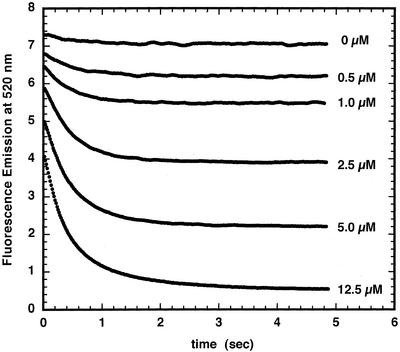
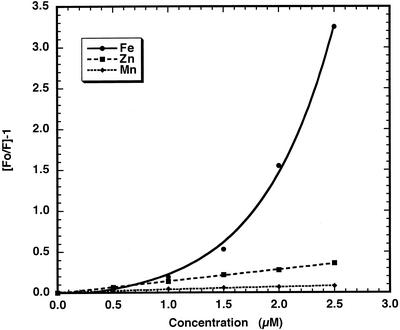
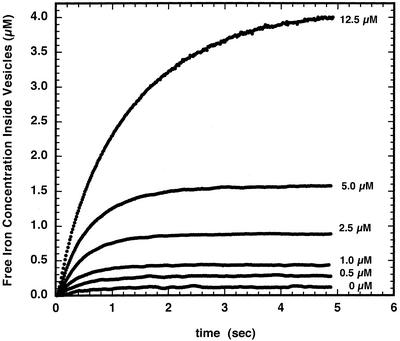
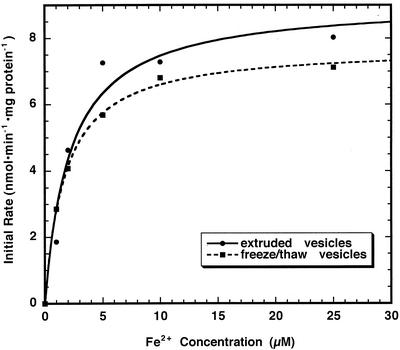
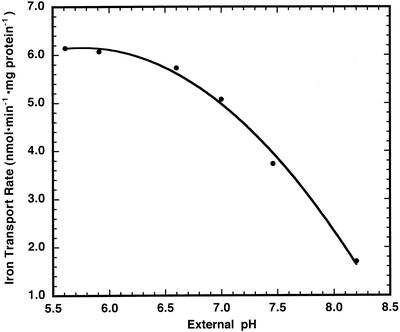
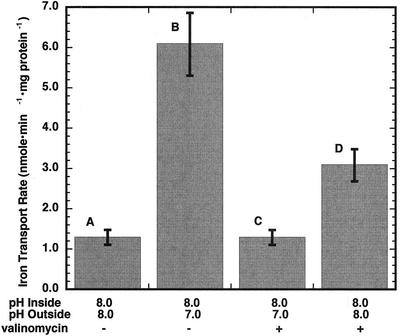
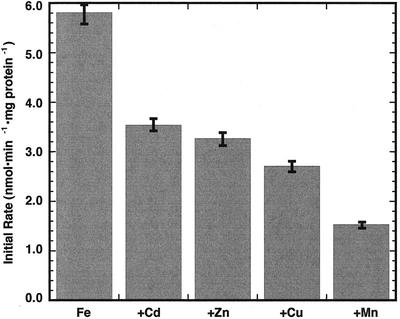
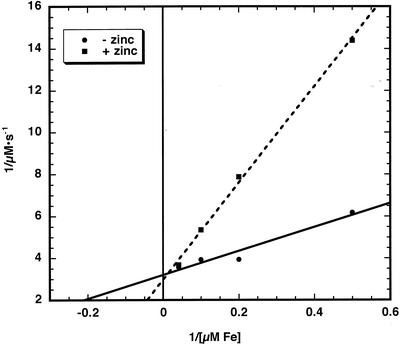
The initial rate of Fe(2+) movement across the inner envelope membrane of pea (Pisum sativum) chloroplasts was directly measured by stopped-flow spectrofluorometry using membrane vesicles loaded with the Fe(2+)-sensitive fluorophore, Phen Green SK. The rate of Fe(2+) transport was rapid, coming to equilibrium within 3s. The maximal rate and concentration dependence of Fe(2+) transport in predominantly right-side-out vesicles were nearly equivalent to those measured in largely inside-out vesicles. Fe(2+) transport was stimulated by an inwardly directed electrochemical proton gradient across right-side-out vesicles, an effect that was diminished by the addition of valinomycin in the presence of K(+). Fe(2+) transport was inhibited by Zn(2+), in a competitive manner, as well as by Cu(2+) and Mn(2+). These results indicate that inward-directed Fe(2+) transport across the chloroplast inner envelope occurs by a potential-stimulated uniport mechanism.
Figures
References
-
- Bensadoun A, Weinstein D. Assay of proteins in the presence of interfering materials. Anal Biochem. 1976;70:241–250. - PubMed
-
- Brüggeman W, Maas-Kantel K, Moog P. Iron uptake by leaf mesophyll cells: the role of the plasma membrane-bound ferric-chelate reductase. Planta. 1993;190:151–155.
-
- Bughio N, Takahashi M, Yoshimura E, Nishizawa NK, Mori S. Light-dependent iron transport into isolated barley chloroplasts. Plant Cell Physiol. 1997;38:101–105.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
