

Genetic variation at the 22q11 PRODH2/DGCR6 locus presents an unusual pattern and increases susceptibility to schizophrenia
- PMID: 11891283
- PMCID: PMC122590
- DOI: 10.1073/pnas.042700699
Genetic variation at the 22q11 PRODH2/DGCR6 locus presents an unusual pattern and increases susceptibility to schizophrenia
Abstract
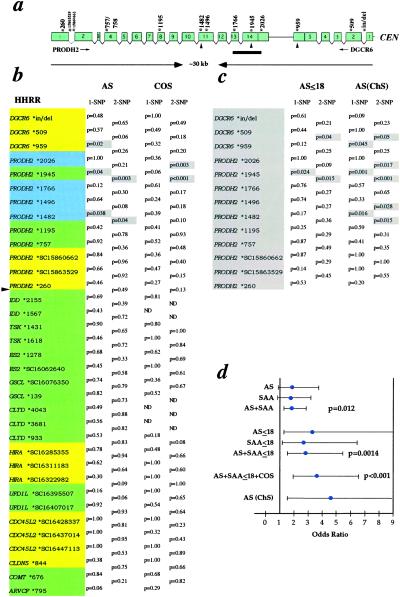
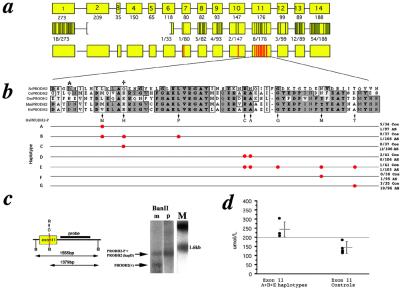
The location of a schizophrenia susceptibility locus at chromosome 22q11 has been suggested by genome-wide linkage studies. Additional support was provided by the observation of a higher-than-expected frequency of 22q11 microdeletions in patients with schizophrenia and the demonstration that approximately 20-30% of individuals with 22q11 microdeletions develop schizophrenia or schizoaffective disorder in adolescence and adulthood. Analysis of the extent of these microdeletions by using polymorphic markers afforded further refinement of this locus to a region of approximately 1.5 Mb. Recently, a high rate of 22q11 microdeletions was also reported for a cohort of 47 patients with Childhood Onset Schizophrenia, a rare and severe form of schizophrenia with onset by age 13. It is therefore likely that this 1.5-Mb region contains one or more genes that predispose to schizophrenia. In three independent samples, we provide evidence for a contribution of the PRODH2/DGCR6 locus in 22q11-associated schizophrenia. We also uncover an unusual pattern of PRODH2 gene variation that mimics the sequence of a linked pseudogene. Several of the pseudogene-like variants we identified result in missense changes at conserved residues and may prevent synthesis of a fully functional enzyme. Our results have implications for understanding the genetic basis of the 22q11-associated psychiatric phenotypes and provide further insights into the genomic instability of this region.
Figures
Comment in
-
A compelling genetic hypothesis for a complex disease: PRODH2/DGCR6 variation leads to schizophrenia susceptibility.Proc Natl Acad Sci U S A. 2002 Apr 16;99(8):4755-6. doi: 10.1073/pnas.092158299. Proc Natl Acad Sci U S A. 2002. PMID: 11959925 Free PMC article. No abstract available.
References
-
- Scambler P J. Hum Mol Genet. 2000;9:2421–2426. - PubMed
-
- Golding-Kushner K J, Weller G, Shprintzen R J. J Cranio-facial Genet Dev Biol. 1985;5:259–266. - PubMed
-
- Pulver A E, Nestadt G, Goldberg R, Shprintzen R J, Lamacz M, Wolyniec P S, Morrow B, Karayiorgou M, Antonarakis S E, Housman D, Kucherlapati R. J Nerv Mental Dis. 1994;182:476–478. - PubMed
-
- Murphy K C, Jones L A, Owen M J. Arch Gen Psychiatry. 1999;56:940–945. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
