

In vitro and in vivo characterization of a Bordetella bronchiseptica mutant strain with a deep rough lipopolysaccharide structure
- PMID: 11895940
- PMCID: PMC127836
- DOI: 10.1128/IAI.70.4.1791-1798.2002
In vitro and in vivo characterization of a Bordetella bronchiseptica mutant strain with a deep rough lipopolysaccharide structure
Abstract
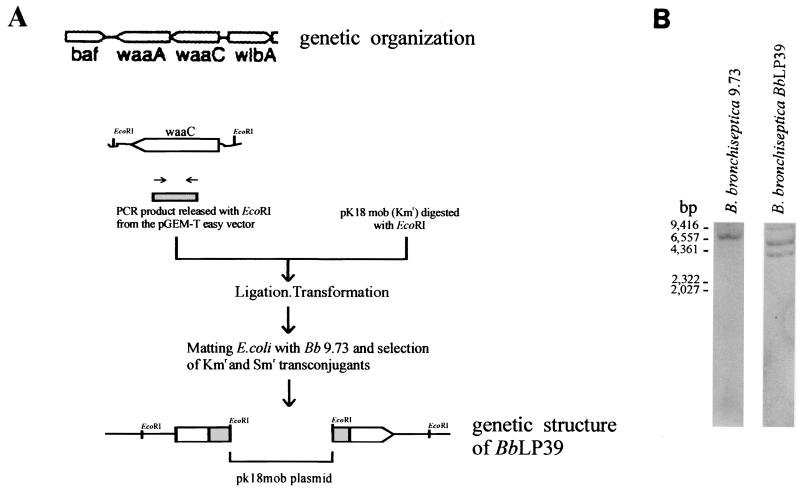
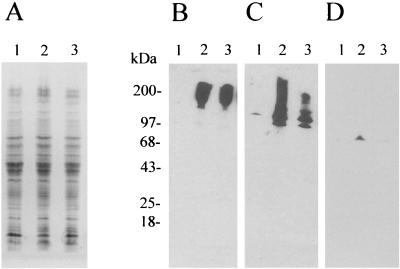
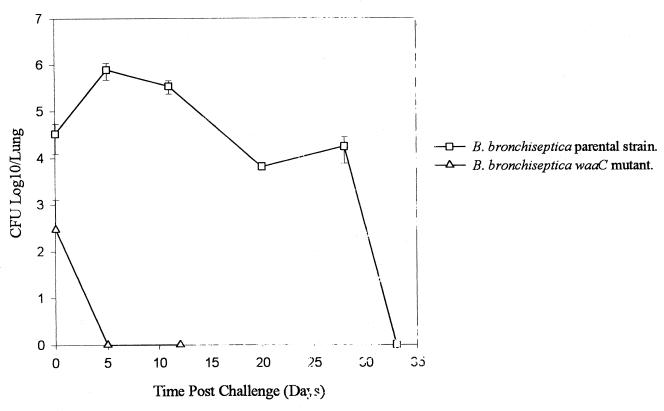
Bordetella bronchiseptica is closely related to Bordetella pertussis, which produces respiratory disease primarily in mammals other than humans. However, its importance as a human pathogen is being increasingly recognized. Although a large amount of research on Bordetella has been generated regarding protein virulence factors, the participation of the surface lipopolysaccharide (LPS) during B. bronchiseptica infection is less understood. To get a better insight into this matter, we constructed and characterized the behavior of an LPS mutant with the deepest possible rough phenotype. We generated the defective mutant B. bronchiseptica LP39 on the waaC gene, which codes for a heptosyl transferase involved in the biosynthesis of the core region of the LPS molecule. Although in B. bronchiseptica LP39 the production of the principal virulence determinants adenylate cyclase-hemolysin, filamentous hemagglutinin, and pertactin persisted, the quantity of the two latter factors was diminished, with the levels of pertactin being the most greatly affected. Furthermore, the LPS of B. bronchiseptica LP39 did not react with sera obtained from mice that had been infected with the parental strain, indicating that this defective LPS is immunologically different from the wild-type LPS. In vivo experiments demonstrated that the ability to colonize the respiratory tract is reduced in the mutant, being effectively cleared from lungs within 5 days, whereas the parental strain survived at least for 30 days. In vitro experiments have demonstrated that, although B. bronchiseptica LP39 was impaired for adhesion to human epithelial cells, it is still able to survive within the host cells as efficiently as the parental strain. These results seem to indicate that the deep rough form of B. bronchiseptica LPS cannot represent a dominant phenotype at the first stage of colonization. Since isolates with deep rough LPS phenotype have already been obtained from human B. bronchiseptica chronic infections, the possibility that this phenotype arises as a consequence of selection pressure within the host at a late stage of the infection process is discussed.
Figures




Similar articles
-
Multiple roles for Bordetella lipopolysaccharide molecules during respiratory tract infection.Infect Immun. 2000 Dec;68(12):6720-8. doi: 10.1128/IAI.68.12.6720-6728.2000. Infect Immun. 2000. PMID: 11083787 Free PMC article.
-
Membrane Vesicles Derived from Bordetella bronchiseptica: Active Constituent of a New Vaccine against Infections Caused by This Pathogen.Appl Environ Microbiol. 2018 Jan 31;84(4):e01877-17. doi: 10.1128/AEM.01877-17. Print 2018 Feb 15. Appl Environ Microbiol. 2018. PMID: 29180369 Free PMC article.
-
A deep rough type structure in Bordetella bronchiseptica lipopolysaccharide modulates host immune responses.Microbiol Immunol. 2011 Dec;55(12):847-54. doi: 10.1111/j.1348-0421.2011.00395.x. Microbiol Immunol. 2011. PMID: 22039958
-
Environmental sensing mechanisms in Bordetella.Adv Microb Physiol. 2001;44:141-81. doi: 10.1016/s0065-2911(01)44013-6. Adv Microb Physiol. 2001. PMID: 11407112 Review.
-
Bordetella bronchiseptica infection of rats and mice.Comp Med. 2003 Feb;53(1):11-20. Comp Med. 2003. PMID: 12625502 Review.
Cited by
-
Homologs of the LapD-LapG c-di-GMP Effector System Control Biofilm Formation by Bordetella bronchiseptica.PLoS One. 2016 Jul 5;11(7):e0158752. doi: 10.1371/journal.pone.0158752. eCollection 2016. PLoS One. 2016. PMID: 27380521 Free PMC article.
-
Cyclic-di-GMP signalling regulates motility and biofilm formation in Bordetella bronchiseptica.Microbiology (Reading). 2013 May;159(Pt 5):869-879. doi: 10.1099/mic.0.064345-0. Epub 2013 Mar 8. Microbiology (Reading). 2013. PMID: 23475948 Free PMC article.
-
Cyclic di-GMP Regulates the Type III Secretion System and Virulence in Bordetella bronchiseptica.Infect Immun. 2022 Jun 16;90(6):e0010722. doi: 10.1128/iai.00107-22. Epub 2022 May 25. Infect Immun. 2022. PMID: 35612302 Free PMC article.
-
Identification of a novel lipopolysaccharide core biosynthesis gene cluster in Bordetella pertussis, and influence of core structure and lipid A glucosamine substitution on endotoxic activity.Infect Immun. 2009 Jul;77(7):2602-11. doi: 10.1128/IAI.00033-09. Epub 2009 Apr 13. Infect Immun. 2009. PMID: 19364841 Free PMC article.
-
Cavitary pneumonia in an AIDS patient caused by an unusual Bordetella bronchiseptica variant producing reduced amounts of pertactin and other major antigens.J Clin Microbiol. 2002 Sep;40(9):3146-54. doi: 10.1128/JCM.40.9.3146-3154.2002. J Clin Microbiol. 2002. PMID: 12202545 Free PMC article.
References
-
- Allen, A., and D. Maskell. 1996. The identification, cloning and mutagenesis of a genetic locus required for lipopolysaccharide biosynthesis in Bordetella pertussis. Mol. Microbiol. 19:37-52. - PubMed
-
- Allen, A. G., R. M. Thomas, J. T. Cadisch, and D. J. Maskell. 1998. Molecular and functional analysis of the lipopolysaccharide biosynthesis locus wlb from Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica. Mol. Microbiol. 29:27-38. - PubMed
-
- Arico, B., V. Scarlato, D. Monack, S. Falkow, and R. Rappuoli. 1991. Structural and genetic analysis of the bvg locus in Bordetella species. Mol. Microbiol. 5:481-491. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
