

Porphyromonas gingivalis lipopolysaccharide is both agonist and antagonist for p38 mitogen-activated protein kinase activation
- PMID: 11895949
- PMCID: PMC127828
- DOI: 10.1128/IAI.70.4.1867-1873.2002
Porphyromonas gingivalis lipopolysaccharide is both agonist and antagonist for p38 mitogen-activated protein kinase activation
Abstract
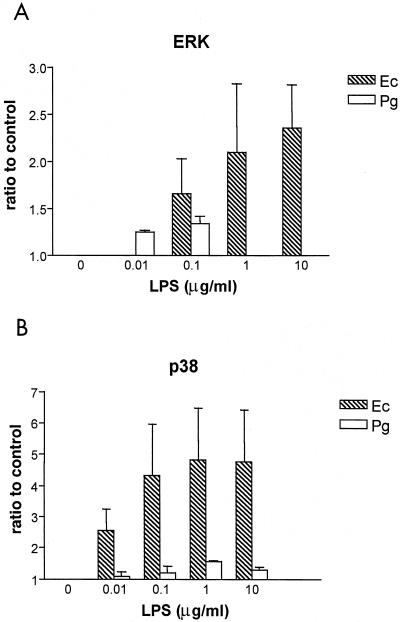
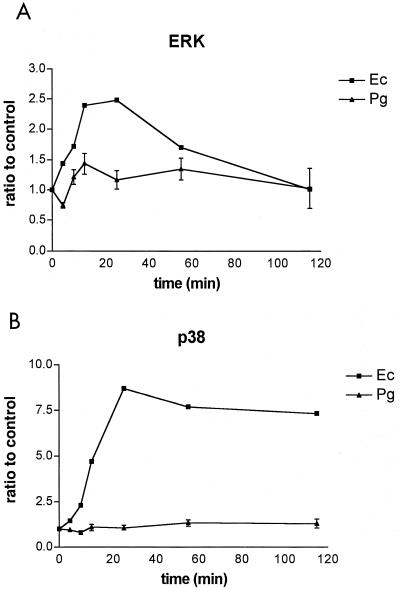
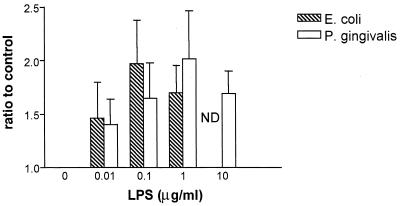
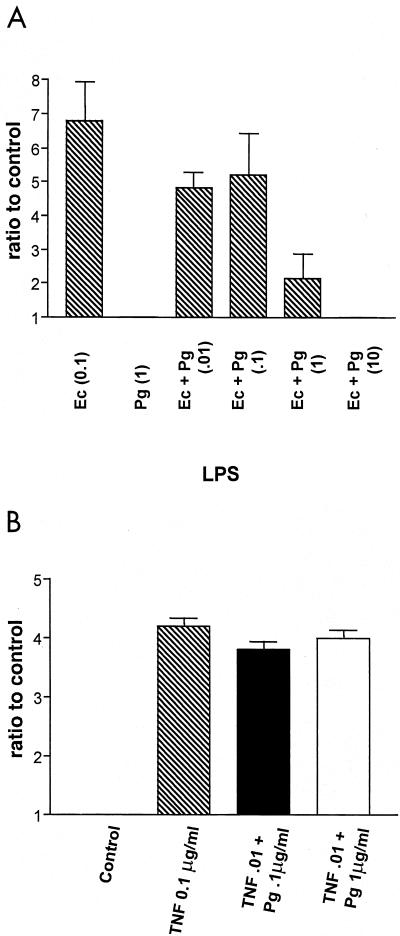
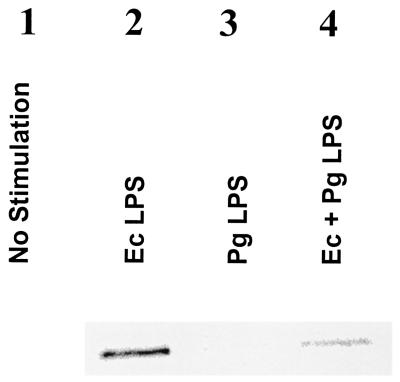
Lipopolysaccharide (LPS) is a key inflammatory mediator. It has been proposed to function as an important molecule that alerts the host of potential bacterial infection. Although highly conserved, LPS contains important structural differences among different bacterial species that can significantly alter host responses. For example, LPS obtained from Porphyromonas gingivalis, an etiologic agent for periodontitis, evokes a highly unusual host cell response. Human monocytes respond to this LPS by the secretion of a variety of different inflammatory mediators, while endothelial cells do not. In addition, P. gingivalis LPS inhibits endothelial cell expression of E-selectin and interleukin 8 (IL-8) induced by other bacteria. In this report the ability of P. gingivalis LPS to activate p38 mitogen-activated protein (MAP) kinase was investigated. It was found that p38 MAP kinase activation occurred in response to P. gingivalis LPS in human monocytes. In contrast, no p38 MAP kinase activation was observed in response to P. gingivalis LPS in human endothelial cells or CHO cells transfected with human Toll-like receptor 4 (TLR-4). In addition, P. gingivalis LPS was an effective inhibitor of Escherichia coli-induced p38 MAP kinase phosphorylation in both endothelial cells and CHO cells transfected with human TLR-4. These data demonstrate that P. gingivalis LPS activates the LPS-associated p38 MAP kinase in monocytes and that it can be an antagonist for E. coli LPS activation of p38 MAP kinase in endothelial and CHO cells. These data also suggest that although LPS is generally considered a bacterial component that alerts the host to infection, LPS from P. gingivalis may selectively modify the host response as a means to facilitate colonization.
Figures
Similar articles
-
Porphyromonas gingivalis lipopolysaccharide: an unusual pattern recognition receptor ligand for the innate host defense system.Acta Odontol Scand. 2001 Jun;59(3):131-8. doi: 10.1080/000163501750266710. Acta Odontol Scand. 2001. PMID: 11501881 Review.
-
Role of the phosphatidylinositol 3 kinase-Akt pathway in the regulation of IL-10 and IL-12 by Porphyromonas gingivalis lipopolysaccharide.J Immunol. 2003 Jul 15;171(2):717-25. doi: 10.4049/jimmunol.171.2.717. J Immunol. 2003. PMID: 12847238
-
Female sex hormones modulate Porphyromonas gingivalis lipopolysaccharide-induced Toll-like receptor signaling in primary human monocytes.J Periodontal Res. 2016 Jun;51(3):395-406. doi: 10.1111/jre.12320. Epub 2015 Sep 14. J Periodontal Res. 2016. PMID: 26364725
-
Differential induction of endotoxin tolerance by lipopolysaccharides derived from Porphyromonas gingivalis and Escherichia coli.J Immunol. 2001 Nov 1;167(9):5278-85. doi: 10.4049/jimmunol.167.9.5278. J Immunol. 2001. PMID: 11673543
-
Porphyromonas gingivalis lipopolysaccharide signaling in gingival fibroblasts-CD14 and Toll-like receptors.Crit Rev Oral Biol Med. 2002;13(2):132-42. doi: 10.1177/154411130201300204. Crit Rev Oral Biol Med. 2002. PMID: 12097356 Review.
Cited by
-
Free lipid A isolated from Porphyromonas gingivalis lipopolysaccharide is contaminated with phosphorylated dihydroceramide lipids: recovery in diseased dental samples.Infect Immun. 2012 Feb;80(2):860-74. doi: 10.1128/IAI.06180-11. Epub 2011 Dec 5. Infect Immun. 2012. PMID: 22144487 Free PMC article.
-
Roles of Porphyromonas gingivalis and its virulence factors in periodontitis.Adv Protein Chem Struct Biol. 2020;120:45-84. doi: 10.1016/bs.apcsb.2019.12.001. Epub 2020 Jan 10. Adv Protein Chem Struct Biol. 2020. PMID: 32085888 Free PMC article. Review.
-
Activation of Notch-1 in oral epithelial cells by P. gingivalis triggers the expression of the antimicrobial protein PLA2-IIA.Mucosal Immunol. 2018 Jul;11(4):1047-1059. doi: 10.1038/s41385-018-0014-7. Epub 2018 Mar 7. Mucosal Immunol. 2018. PMID: 29515164 Free PMC article.
-
Enhancing attachment of human gingival fibroblasts to periodontally compromised teeth: A comparative analysis of hyaluronic acid, EDTA, enamel matrix derivatives.Saudi Dent J. 2023 Jul;35(5):547-552. doi: 10.1016/j.sdentj.2023.05.005. Epub 2023 May 12. Saudi Dent J. 2023. PMID: 37520595 Free PMC article.
-
Hemoglobin receptor protein from Porphyromonas gingivalis induces interleukin-8 production in human gingival epithelial cells through stimulation of the mitogen-activated protein kinase and NF-κB signal transduction pathways.Infect Immun. 2014 Jan;82(1):202-11. doi: 10.1128/IAI.01140-12. Epub 2013 Oct 14. Infect Immun. 2014. PMID: 24126532 Free PMC article.
References
-
- Ajizian, S. J., B. K. English, and E. A. Meals. 1999. Specific inhibitors of p38 and extracellular signal-regulated kinase mitogen-activated protein kinase pathways block inducible nitric oxide synthase and tumor necrosis factor accumulation in murine macrophages stimulated with lipopolysaccharide and interferon-gamma. J. Infect. Dis. 179:939-944. - PubMed
-
- Arbabi, S., M. R. Rosengart, I. Garcia, S. Jelacic, and R. V. Maier. 1999. Priming interleukin 8 production: role of platelet-activating factor and p38. Arch. Surg. 134:1348-1353. - PubMed
-
- Arbibe, L., J. P. Mira, N. Teusch, L. Kline, M. Guha, N. Mackman, P. J. Godowski, R. J. Ulevitch, and U. G. Knaus. 2000. Toll-like receptor 2-mediated NF-kB activation requires a Rac1-dependent pathway. Nat. Immunol. 1:533-540. - PubMed
-
- Arditi, M., J. Zhou, M. Torres, D. L. Durden, M. Stins, and K. S. Kim. 1995. Lipopolysaccharide stimulates the tyrosine phosphorylation of mitogen-activated protein kinases p44, p42, and p41 in vascular endothelial cells in a soluble CD14-dependent manner. Role of protein tyrosine phosphorylation in lipopolysaccharide-induced stimulation of endothelial cells. J. Immunol. 155:3994-4003. - PubMed
-
- Bainbridge, B. W., and R. P. Darveau. 1997. Lipopolysaccharide from oral bacteria: role in innate host defense and chronic inflammatory disease, p. 899-913. In D. Morrison (ed.), Endotoxin in health and disease. Marcel Dekker, New York, N.Y.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
