

Long-lasting potentiation of GABAergic synapses in dopamine neurons after a single in vivo ethanol exposure
- PMID: 11896147
- PMCID: PMC6758244
- DOI: 10.1523/JNEUROSCI.22-06-02074.2002
Long-lasting potentiation of GABAergic synapses in dopamine neurons after a single in vivo ethanol exposure
Abstract
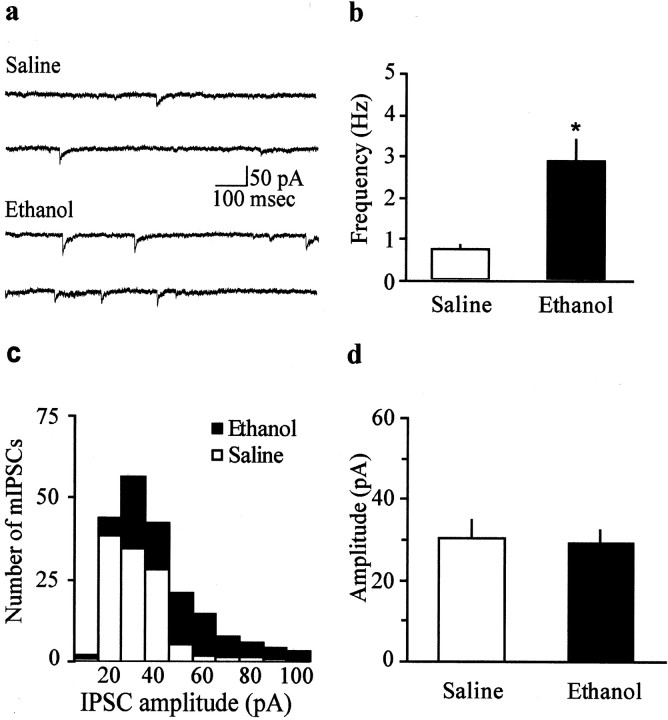
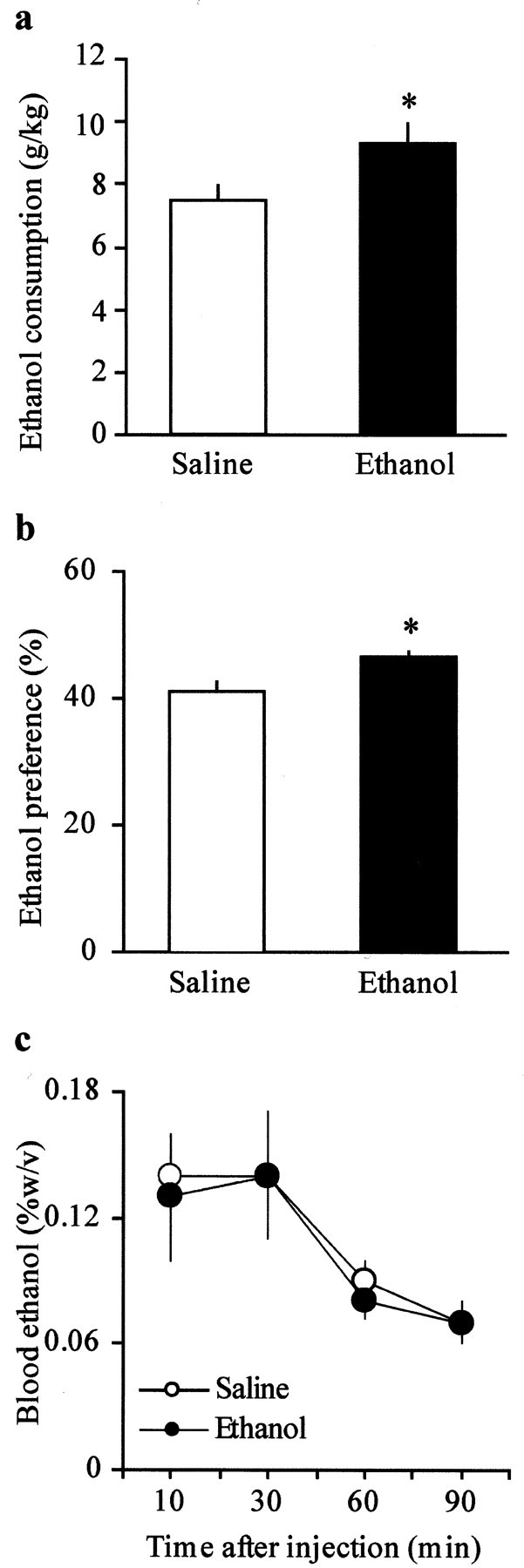
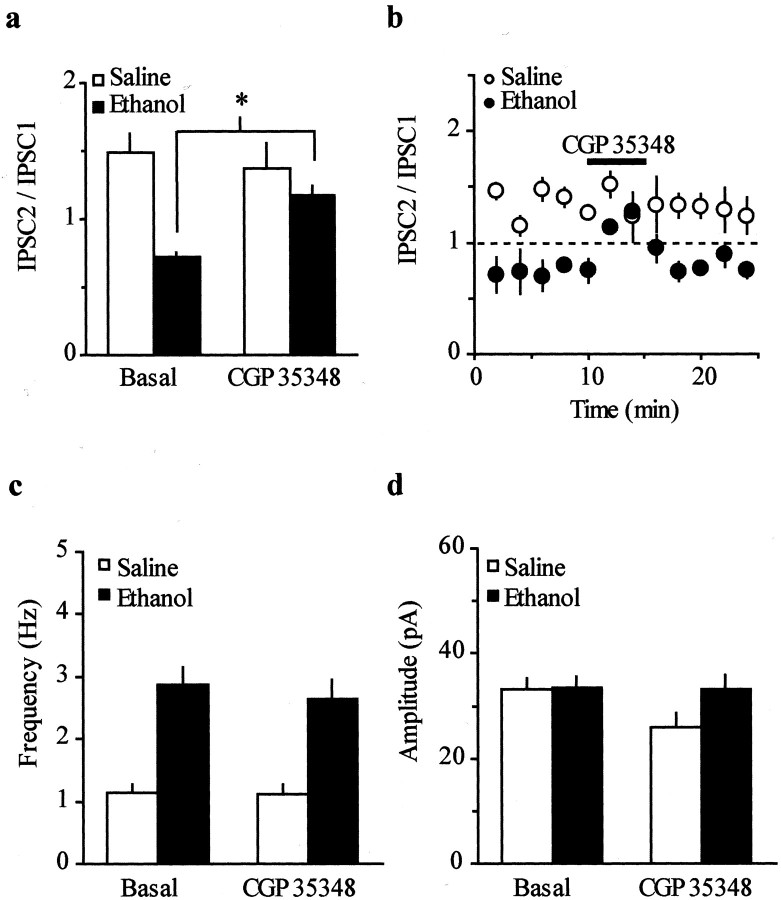
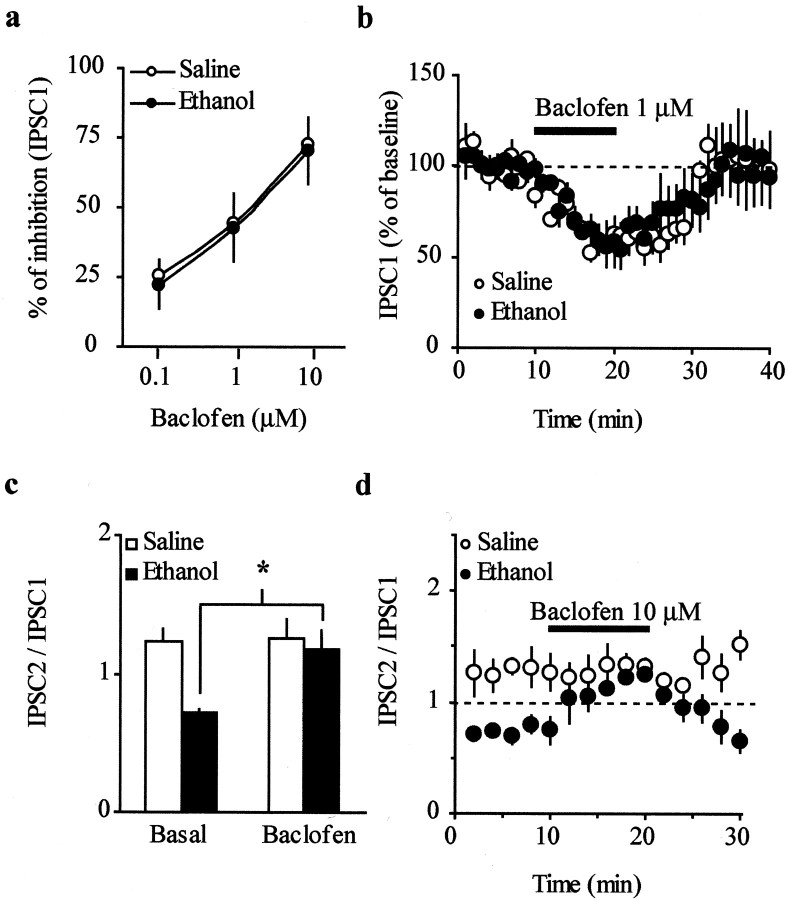
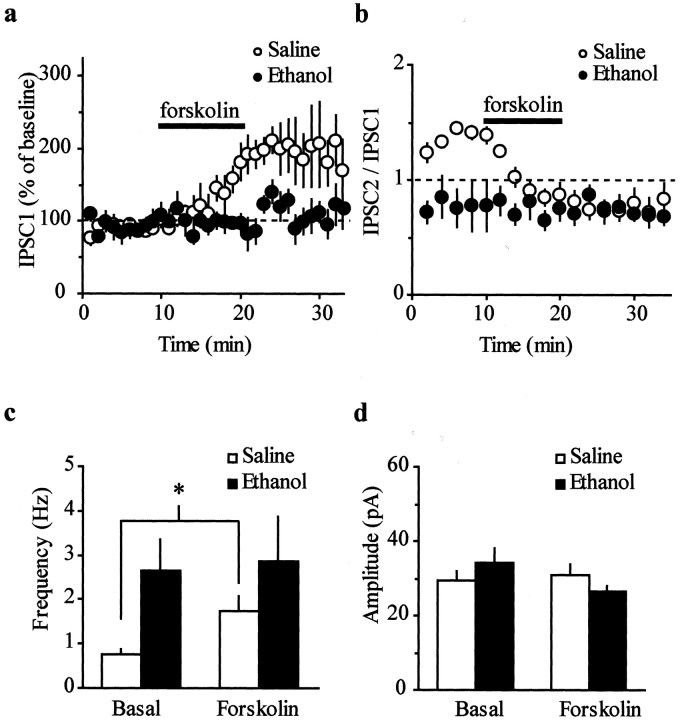
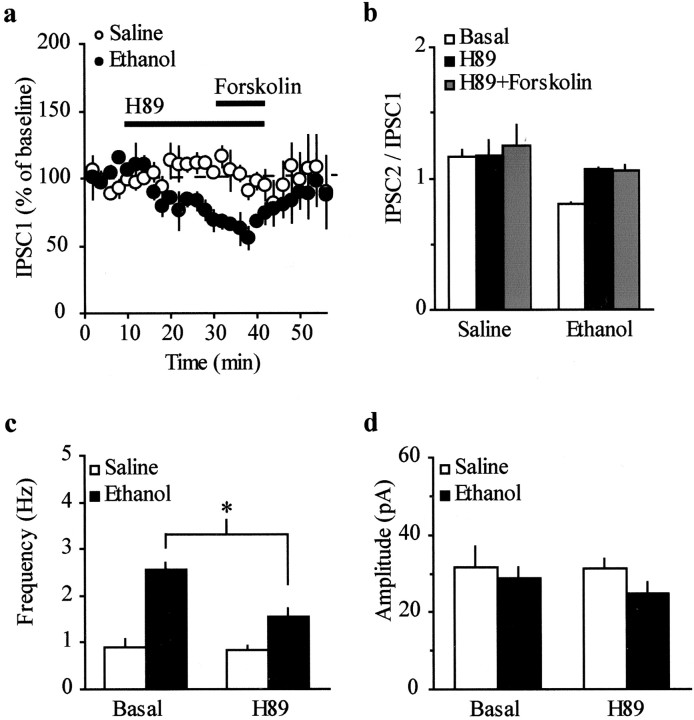
The mesolimbic dopamine (DA) system originating in the ventral tegmental area (VTA) is involved in many drug-related behaviors, including ethanol self-administration. In particular, VTA activity regulating ethanol consummatory behavior appears to be modulated through GABA(A) receptors. Previous exposure to ethanol enhances ethanol self-administration, but the mechanisms underlying this phenomenon are not well understood. In this study, we examined changes occurring at GABA synapses onto VTA DA neurons after a single in vivo exposure to ethanol. We observed that evoked GABA(A) IPSCs in DA neurons of ethanol-treated animals exhibited paired-pulse depression (PPD) compared with saline-treated animals, which exhibited paired-pulse facilitation (PPF). Furthermore, PPD was still present 1 week after the single exposure to ethanol. An increase in frequency of spontaneous miniature GABA(A) IPSCs (mIPSCs) was also observed in the ethanol-treated animals. Additionally, the GABA(B) receptor antagonist (3-aminopropyl)(diethoxymethyl) phosphinic acid shifted PPD to PPF, indicating that presynaptic GABA(B) receptor activation, likely attributable to GABA spillover, might play a role in mediating PPD in the ethanol-treated mice. The activation of adenylyl cyclase by forskolin increased the amplitude of GABA(A) IPSCs and the frequency of mIPSCs in the saline- but not in the ethanol-treated animals. Conversely, the protein kinase A (PKA) inhibitor N-[z-(p-bromocinnamylamino)ethyl]-5-isoquinolinesulfonamide significantly decreased both the frequency of spontaneous mIPSCs and the amplitude of GABA(A) IPSCs in the ethanol-treated mice but not in the saline controls. The present results indicate that potentiation of GABAergic synapses, via a PKA-dependent mechanism, occurs in the VTA after a single in vivo exposure to ethanol, and such potentiation might be a key synaptic modification underlying increased ethanol intake.
Figures

References
-
- Aguayo LG, Pancetti FC. Ethanol modulation of the gamma-aminobutyric acidA- and glycine-activated Cl− current in cultured mouse neurons. J Pharmacol Exp Ther. 1994;270:61–69. - PubMed
-
- Andretic R, Chaney S, Hirsh J. Requirement of circadian genes for cocaine sensitization in Drosophila. Science. 1999;285:1066–1068. - PubMed
-
- Bailey CP, Manley SJ, Watson WP, Wonnacott S, Molleman A, Little HJ. Chronic ethanol administration alters activity in ventral tegmental area neurons after cessation of withdrawal hyperexcitability. Brain Res. 1998;803:144–152. - PubMed
-
- Boyle AE, Segal R, Smith BR, Amit Z. Bidirectional effects of GABAergic agonists and antagonists on maintenance of voluntary ethanol intake in laboratory rats. Pharmacol Biochem Behav. 1993;46:179–182. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources