

Elevated fusiform cell activity in the dorsal cochlear nucleus of chinchillas with psychophysical evidence of tinnitus
- PMID: 11896177
- PMCID: PMC6758251
- DOI: 10.1523/JNEUROSCI.22-06-02383.2002
Elevated fusiform cell activity in the dorsal cochlear nucleus of chinchillas with psychophysical evidence of tinnitus
Abstract
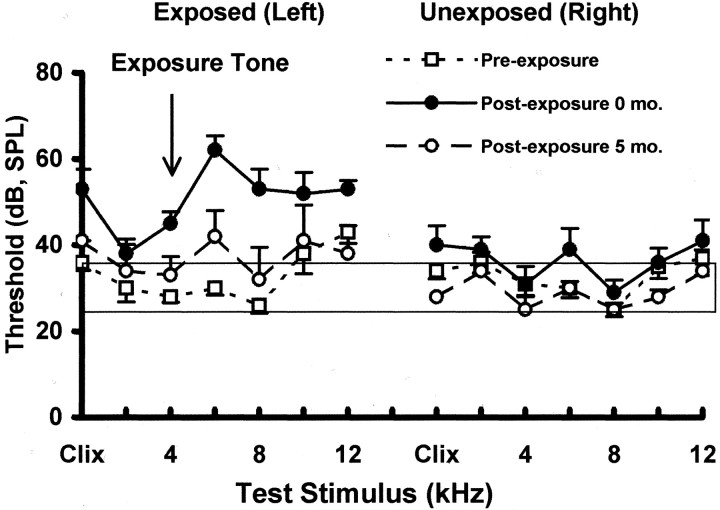
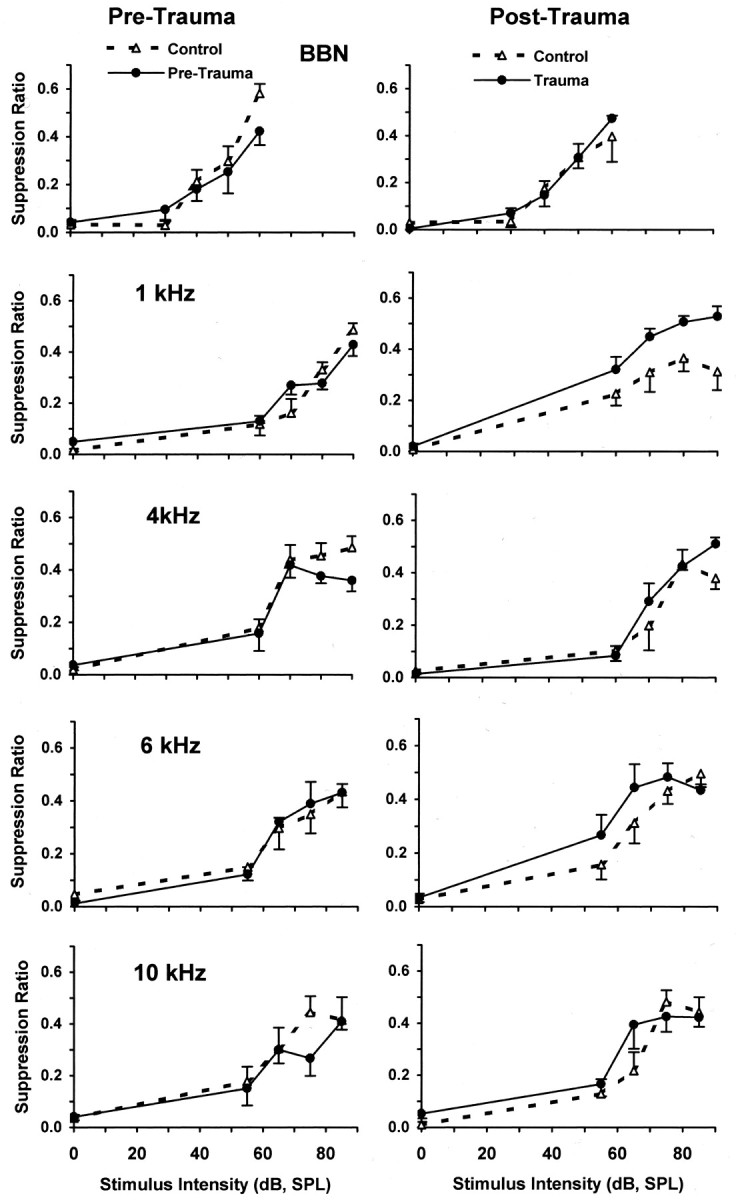
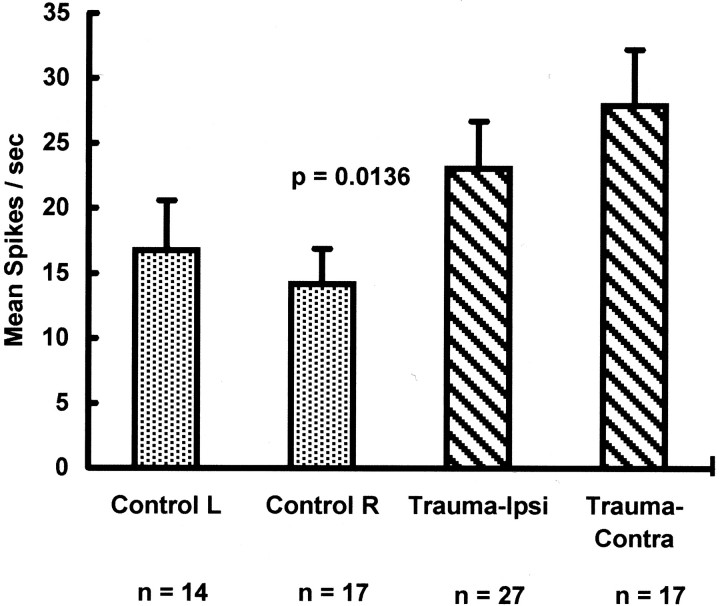
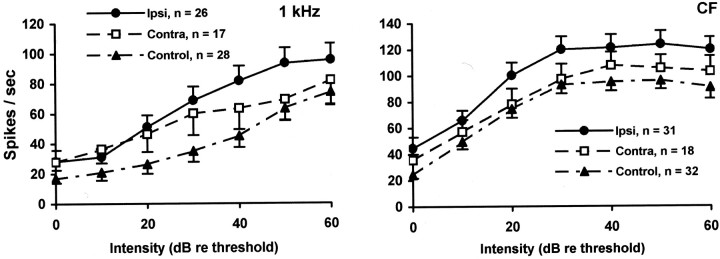
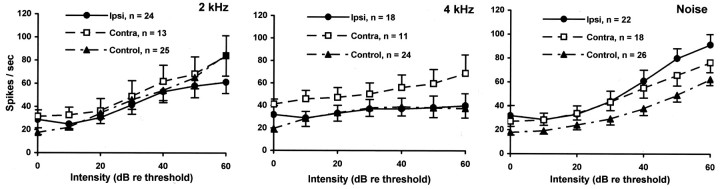
Chinchillas with psychophysical evidence of chronic tinnitus were shown to have significantly elevated spontaneous activity and stimulus-evoked responses in putative fusiform cells of the dorsal cochlear nuclei (DCN). Chinchillas were psychophysically trained and tested before and after exposure to a traumatic unilateral 80 dB (sound pressure level) 4 kHz tone. Before exposure, two groups were matched in terms of auditory discrimination performance (noise, and 1, 4, 6, and 10 kHz tones). After exposure, a single psychophysical difference emerged between groups. The exposed group displayed enhanced discrimination of 1 kHz tones (p = 0.00027). Postexposure discrimination of other stimuli was unaffected. It was hypothesized that exposed animals experienced a chronic subjective tone (i.e., tinnitus), resulting from their trauma, and that features of this subjective tone were similar enough to 1 kHz to affect discrimination of 1 kHz objective signals. After psychophysical testing, single-unit recordings were obtained from each animal's DCN fusiform cell layer. Putative fusiform cells of exposed animals showed significantly (p = 0.0136) elevated spontaneous activity, compared with cells of unexposed animals. Putative fusiform cells of exposed animals showed a greater stimulus-evoked response to tones at 1 kHz (p = 0.0000006) and at characteristic-frequency (p = 0.0000009). This increased activity was more pronounced on the exposed side. No increase in stimulus-evoked responses was observed to other frequencies or noise. These parallel psychophysical and electrophysiological results are consistent with the hypothesis that chronic tonal tinnitus is associated with, and may result from, trauma-induced elevation of activity of DCN fusiform cells.
Figures

References
-
- Alibardi L. Cytology of large neurons in the guinea pig dorsal cochlear nucleus contacting the inferior colliculus. Eur J Histochem. 2000;44:365–375. - PubMed
-
- Anari M, Axelsson A, Eliasson A, Magnusson L. Hypersensitivity to sound–questionnaire data, audiometry and classification. Scand Audiol. 1999;28:219–230. - PubMed
-
- Atherley GRC, Hempstock TI, Noble WG. Study of tinnitus induced temporarily by noise. J Acoust Soc Am. 1968;44:1503–1506. - PubMed
-
- Backoff PM, Palombi PS, Caspary DM. Glycinergic and GABAergic inputs affect short-term suppression in the cochlear nucleus. Hear Res. 1997;110:155–163. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous