

Polycomb group gene rae28 is required for sustaining activity of hematopoietic stem cells
- PMID: 11901201
- PMCID: PMC2193743
- DOI: 10.1084/jem.20011911
Polycomb group gene rae28 is required for sustaining activity of hematopoietic stem cells
Abstract
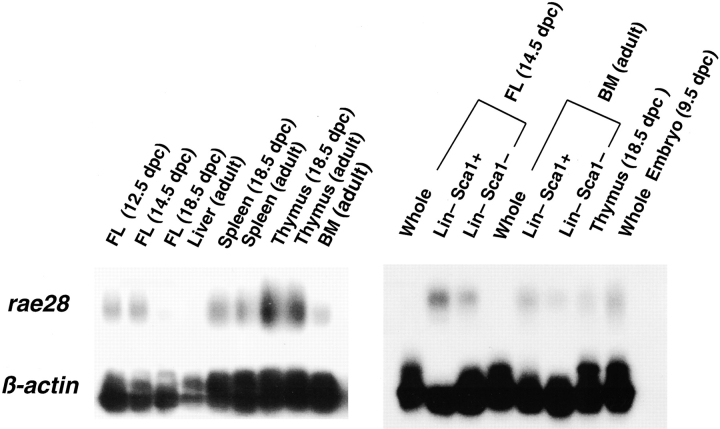
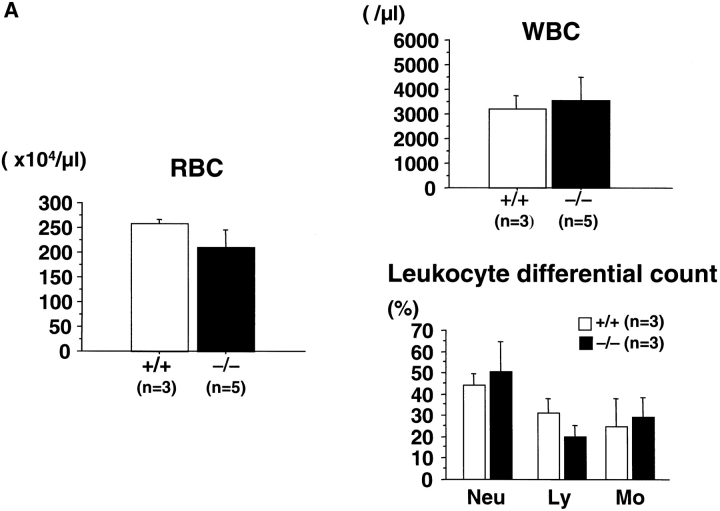
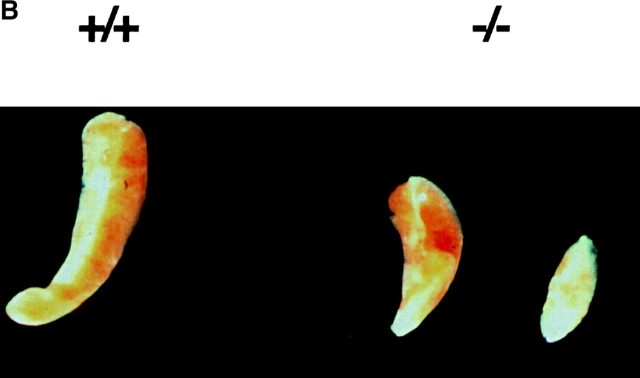
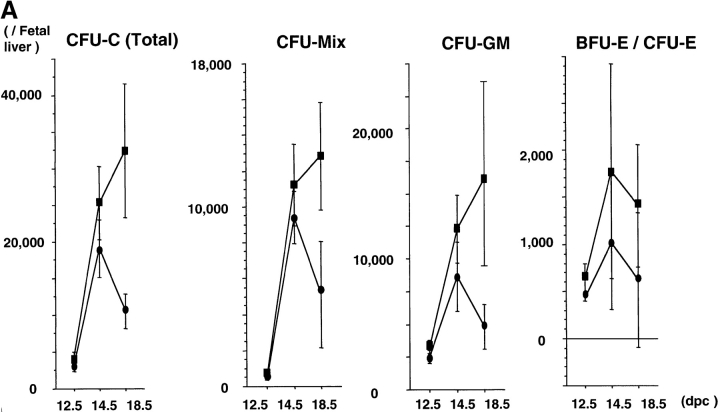
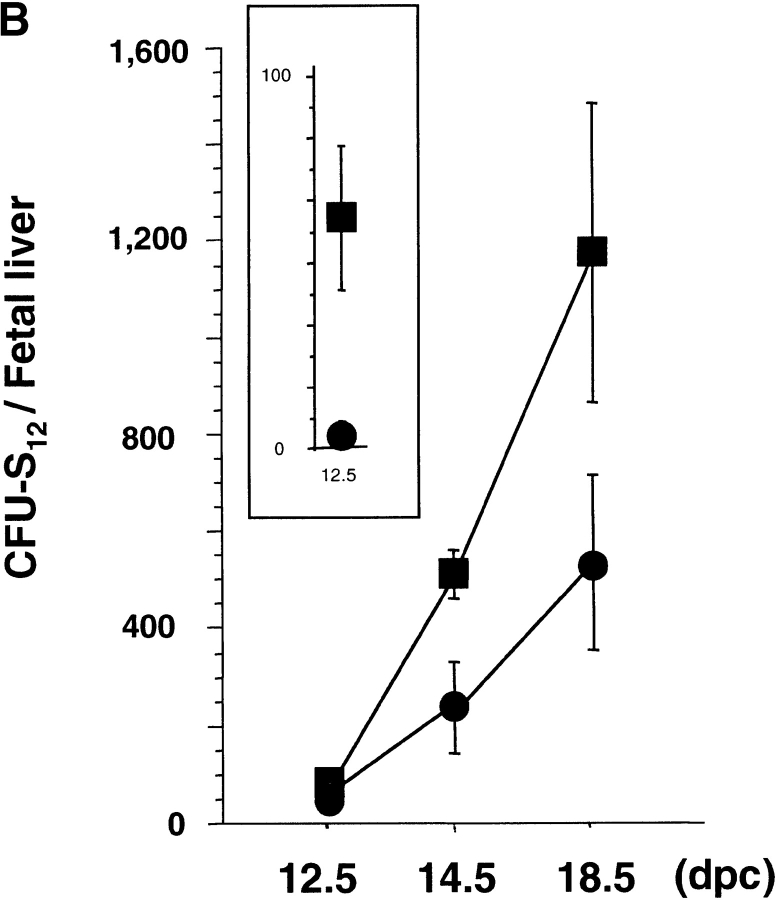
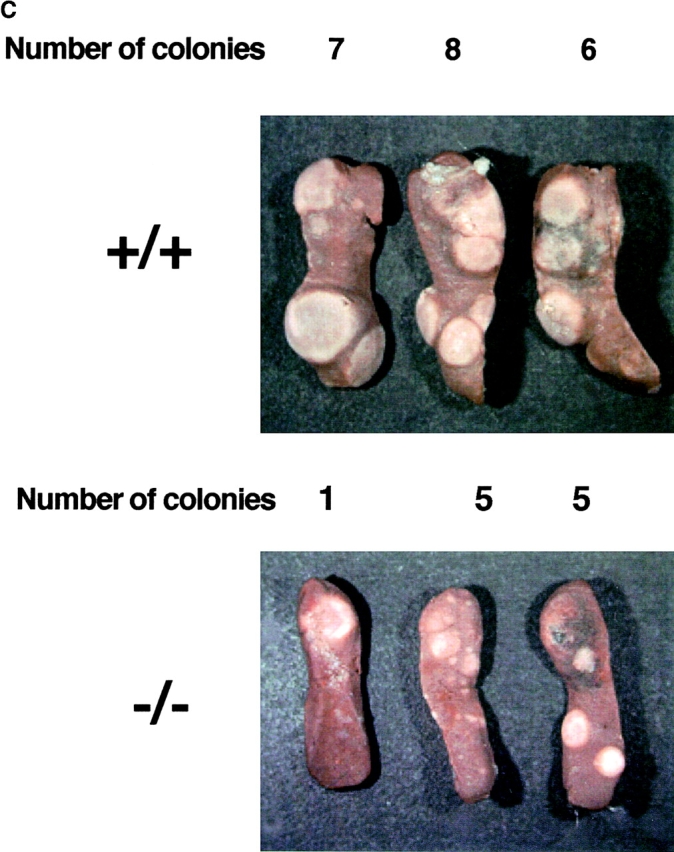
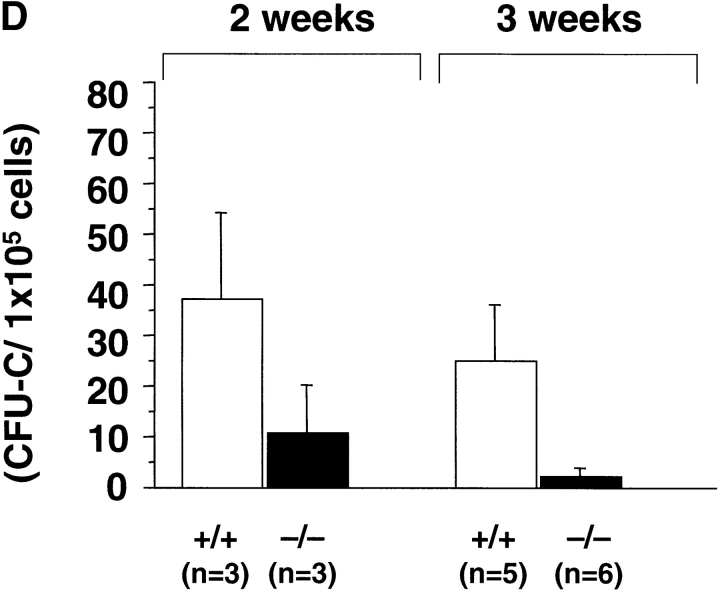
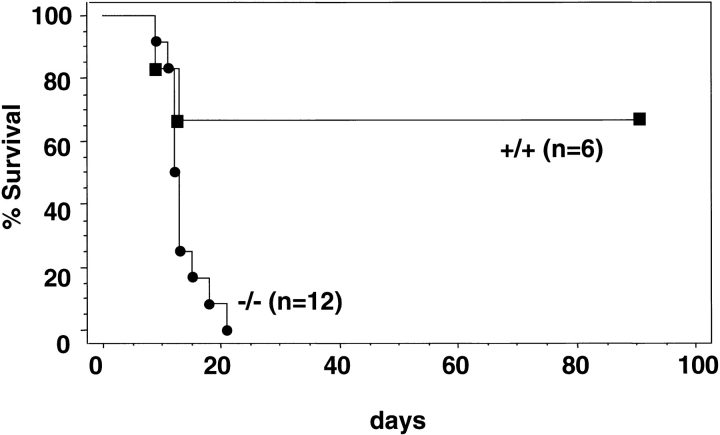
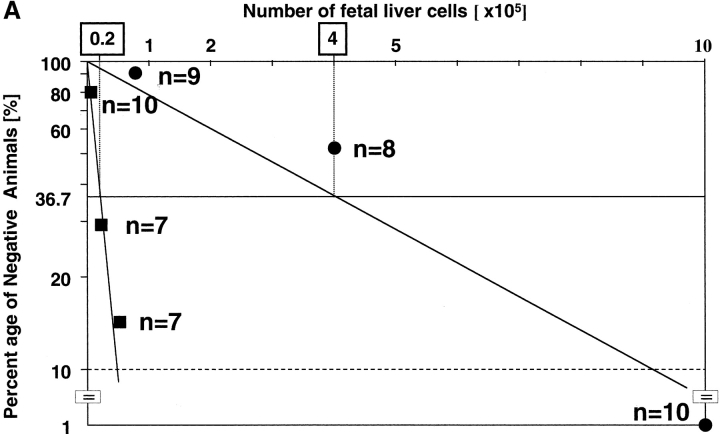
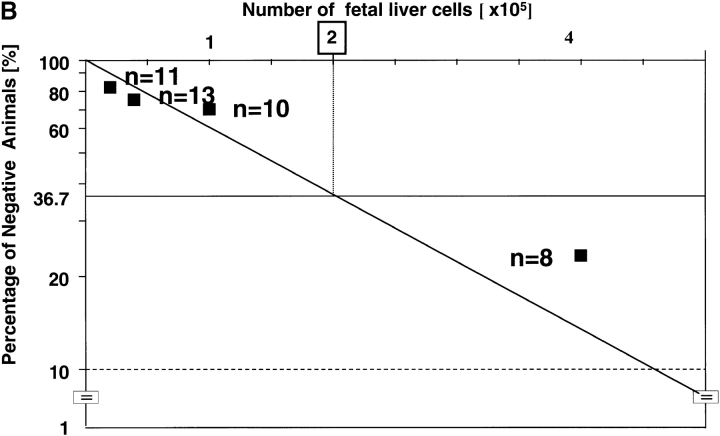
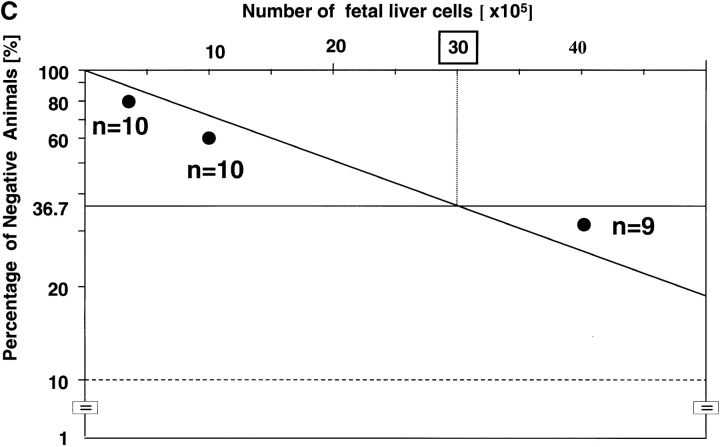
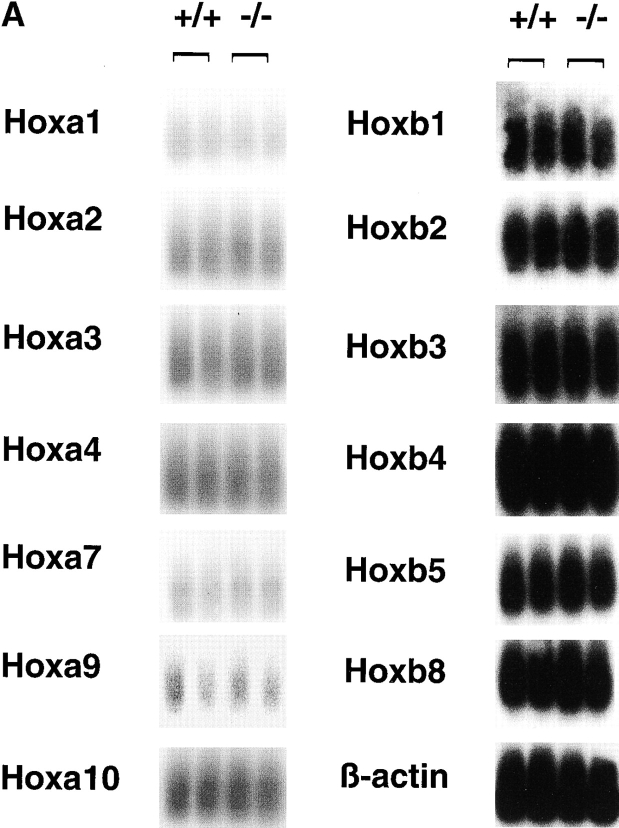
The rae28 gene (rae28), also designated as mph1, is a mammalian ortholog of the Drosophila polyhomeotic gene, a member of Polycomb group genes (PcG). rae28 constitutes PcG complex 1 for maintaining transcriptional states which have been once initiated, presumably through modulation of the chromatin structure. Hematopoietic activity was impaired in the fetal liver of rae28-deficient animals (rae28-/-), as demonstrated by progressive reduction of hematopoietic progenitors of multilineages and poor expansion of colony forming units in spleen (CFU-S(12)) during embryonic development. An in vitro long-term culture-initiating cell assay suggested a reduction in hematopoietic stem cells (HSCs), which was confirmed in vivo by reconstitution experiments in lethally irradiated congenic recipient mice. The competitive repopulating units (CRUs) reflect HSCs supporting multilineage blood-cell production. CRUs were generated, whereas the number of CRUs was reduced by a factor of 20 in the rae28-/- fetal liver. We also performed serial transplantation experiments to semiquantitatively measure self-renewal activity of CRUs in vivo. Self-renewal activity of CRUs was 15-fold decreased in rae28-/-. Thus the compromised HSCs were presumed to reduce hematopoietic activity in the rae28-/- fetal liver. This is the first report to suggest that rae28 has a crucial role in sustaining the activity of HSCs to maintain hematopoiesis.
Figures

References
-
- Simon, J. 1995. Locking in stable states of gene expression: transcriptional control during Drosophila development. Curr. Opin. Cell Biol. 7:376–385. - PubMed
-
- Gould, A. 1997. Functions of mammalian Polycomb group and trithorax group related genes. Curr. Opin. Genet. Dev. 7:488–494. - PubMed
-
- Brock, H.W., and M. van Lohuizen. 2001. The polycomb group-no longer an exclusive club? Curr. Opin. Genet. Dev. 11:175–181. - PubMed
-
- Nomura, M., Y. Takihara, and K. Shimada. 1994. Isolation and characterization of retinoic acid-inducible cDNA clones in F9 cells: one of the early inducible clones encodes a novel protein sharing several highly homologous regions with a Drosophila polyhomeotic protein. Differentiation. 57:39–50. - PubMed
-
- Ohta, H., S. Tokimasa, Z. Zou, S. Funaki, H. Kurahashi, Y. Takahashi, M. Kimura, R. Matsuoka, M. Horie, J. Hara, et al. 2000. Structure and chromosomal localization of the RAE28/HPH1 gene, a human homologue of the polyhomeotic gene. DNA Seq. 11:61–73. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
