

Parameters of human hepatitis delta virus genome replication: the quantity, quality, and intracellular distribution of viral proteins and RNA
- PMID: 11907210
- PMCID: PMC136113
- DOI: 10.1128/jvi.76.8.3709-3719.2002
Parameters of human hepatitis delta virus genome replication: the quantity, quality, and intracellular distribution of viral proteins and RNA
Abstract
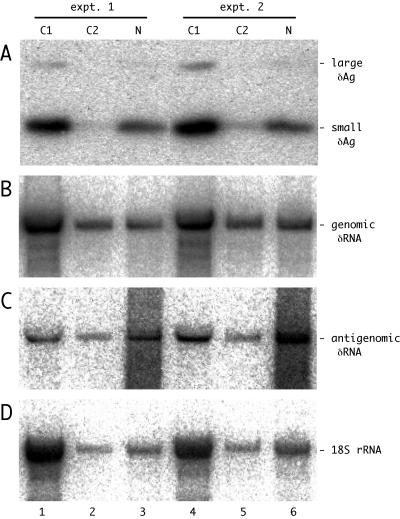
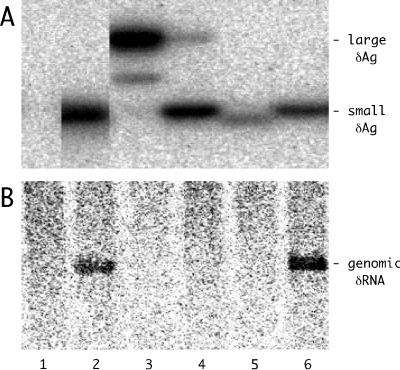
Assembly of hepatitis delta virus (HDV) in infected human hepatocytes involves association of the 1,679- nucleotide single-stranded genomic RNA (deltaRNA) with multiple copies of both small and large forms of the delta protein (deltaAg) to form a ribonucleoprotein particle which in turn interacts with envelope proteins of the natural helper virus, hepatitis B virus. Subsequently, for initiation of a new round of replication, the amount of small deltaAg within the assembled HDV particle is both necessary and sufficient. Quantitative assays were used in order to better understand just how much deltaAg is needed. The molar ratio of deltaAg species to genomic deltaRNA in assembled HDV particles was approximately 200. Next, this ratio was determined for cells under several different experimental situations in which HDV genome replication was occurring. These included replication in woodchuck liver and also in mouse liver and skeletal muscle, as well as replication in stably and transiently transfected cultured human hepatoblastoma cells. Surprisingly, in almost all these situations the molar ratios were comparable to that observed for HDV particles. This was true for different times after the initiation of replication and was independent of whether or not virus assembly was occurring. Cell fractionation combined with quantitative assays was used to test whether the majority of deltaAg and deltaRNA were colocalized during HDV replication in transfected cells. The cytoplasmic fraction contained the majority of deltaAg and genomic deltaRNA. Finally, the quality of deltaAg and deltaRNA, especially at relatively late times after the initiation of replication, was examined by using reverse transcription-PCR, cloning, and sequencing through the entire deltaAg open reading frame. When virus assembly and spread were not possible, 20% or less of the predicted deltaAg would have been able to support HDV replication. In summary, an examination of the quantity, quality and intracellular distribution of deltaAg and deltaRNA in several different experimental systems has provided a better understanding of the parameters associated with the initiation, maintenance, and ultimate decline of HDV genome replication.
Figures




References
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. A. Smith, J. G. Seidman, and K. Struhl. 1987. Current protocols in molecular biology. Wiley, New York, N.Y.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
