

Membrane recognition by vesicular stomatitis virus involves enthalpy-driven protein-lipid interactions
- PMID: 11907215
- PMCID: PMC136106
- DOI: 10.1128/jvi.76.8.3756-3764.2002
Membrane recognition by vesicular stomatitis virus involves enthalpy-driven protein-lipid interactions
Abstract
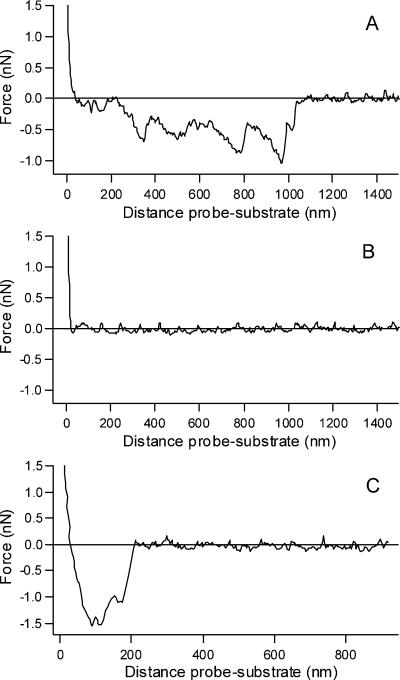
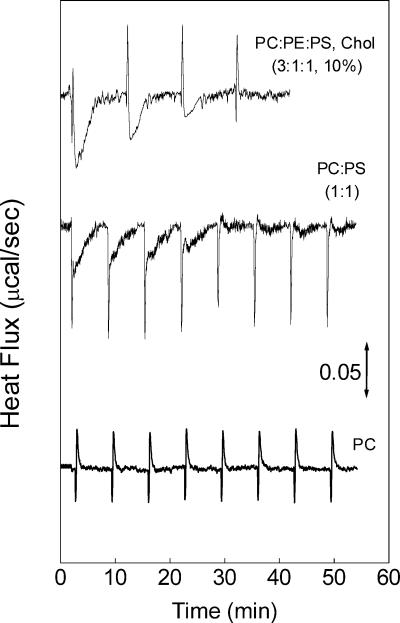
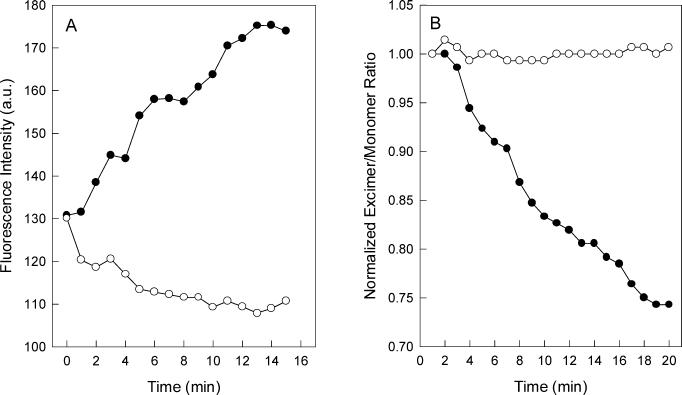
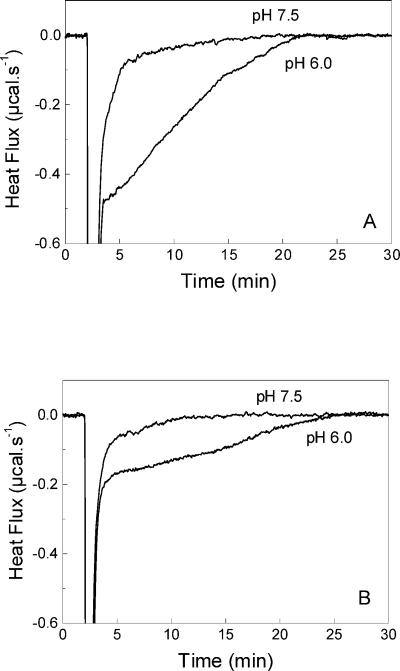
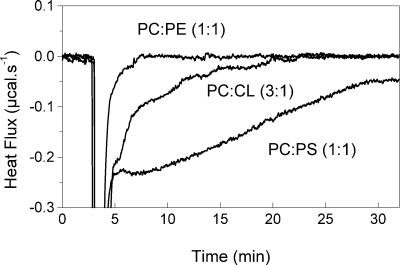
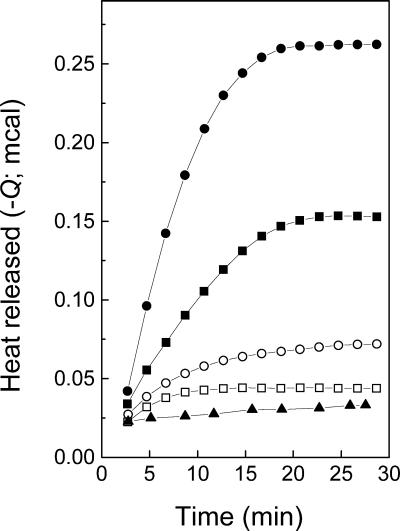
Vesicular stomatitis virus (VSV) infection depends on the fusion of viral and cellular membranes, which is mediated by virus spike glycoprotein G at the acidic environment of the endosomal compartment. VSV G protein does not contain a hydrophobic amino acid sequence similar to the fusion peptides found among other viral glycoproteins, suggesting that membrane recognition occurs through an alternative mechanism. Here we studied the interaction between VSV G protein and liposomes of different phospholipid composition by force spectroscopy, isothermal titration calorimetry (ITC), and fluorescence spectroscopy. Force spectroscopy experiments revealed the requirement for negatively charged phospholipids for VSV binding to membranes, suggesting that this interaction is electrostatic in nature. In addition, ITC experiments showed that VSV binding to liposomes is an enthalpically driven process. Fluorescence data also showed the lack of VSV interaction with the vesicles as well as inhibition of VSV-induced membrane fusion at high ionic strength. Intrinsic fluorescence measurements showed that the extent of G protein conformational changes depends on the presence of phosphatidylserine (PS) on the target membrane. Although the increase in PS content did not change the binding profile, the rate of the fusion reaction was remarkably increased when the PS content was increased from 25 to 75%. On the basis of these data, we suggest that G protein binding to the target membrane essentially depends on electrostatic interactions, probably between positive charges on the protein surface and negatively charged phospholipids in the cellular membrane. In addition, the fusion is exothermic, indicating no entropic constraints to this process.
Figures
References
-
- Bailey, C. A., D. K. Miller, and J. Lenard. 1984. Effects of DEAE-dextran on infection and hemolysis by VSV. Evidence that nonspecific electrostatic interactions mediate effective binding of VSV to cells. Virology 133:111-117. - PubMed
-
- Baranowski, E., C. M. Ruiz-Jarabo, and E. Domingo. 2001. Evolution of cell recognition by viruses. Science 292:1102-1105. - PubMed
-
- Bonnafous, P., and T. Stegmann. 2000. Membrane perturbation and fusion pore formation in influenza hemagglutinin-mediated membrane fusion. A new model for fusion. J. Biol. Chem. 275:6160-6166. - PubMed
-
- Bullough, P. A., F. M. Hughson, J. J. Skehel, and D. C. Wiley. 1994. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature 371:37-43. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
