

MSK1 and MSK2 are required for the mitogen- and stress-induced phosphorylation of CREB and ATF1 in fibroblasts
- PMID: 11909979
- PMCID: PMC133730
- DOI: 10.1128/MCB.22.8.2871-2881.2002
MSK1 and MSK2 are required for the mitogen- and stress-induced phosphorylation of CREB and ATF1 in fibroblasts
Abstract
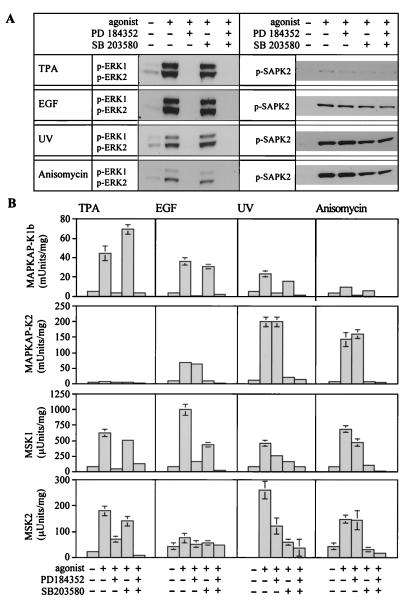
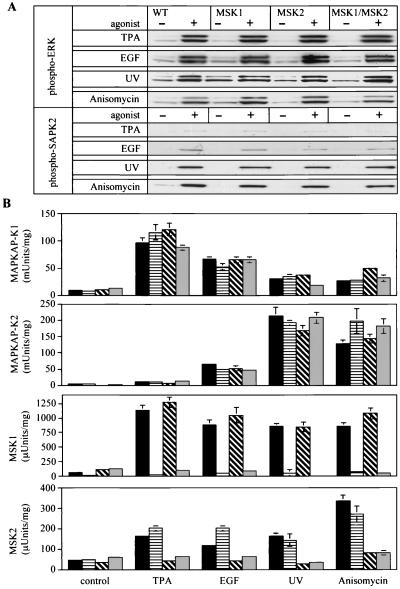
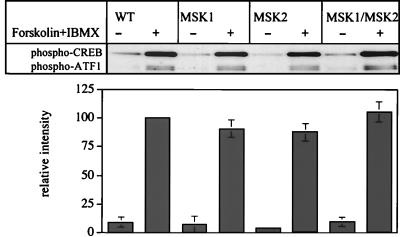
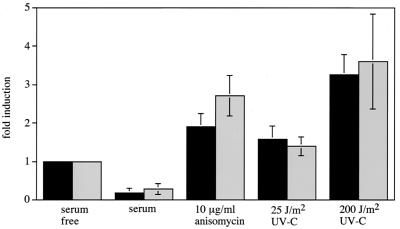
Using mouse knockouts for mitogen- and stress-activated protein kinase 1 (MSK1) and MSK2 and a double knockout of both MSK1 and MSK2, we show that these protein kinases are required for the stress-induced phosphorylation of transcription factors CREB and ATF1 in primary embryonic fibroblasts. In contrast mitogen-induced phosphorylation of CREB and ATF1 is greatly reduced but not totally abolished. The mitogen- and stress-induced phosphorylation of CREB at Ser133 has been linked to the transcription of several immediate early genes, including c-fos, junB, and egr1. The knockout of both MSK1 and MSK2 resulted in a 50% reduction in c-fos and junB gene transcription in response to anisomycin or UV-C radiation but only a small reduction in response to tetradecanoyl phorbol acetate or epidermal growth factor in fibroblasts. The transcription of egr1 in response to both mitogenic and stress stimuli, as well as stress-induced apoptosis, was unaffected in the MSK1/MSK2 double knockout.
Figures








References
-
- Alessi, D. R., A. Cuenda, P. Cohen, D. T. Dudley, and A. R. Saltiel. 1995. PD 098059 is a specific inhibitor of the activation of mitogen-activated protein kinase kinase in vitro and in vivo. J. Biol. Chem. 270:27489-27494. - PubMed
-
- Amato, S. F., K. Nakajima, T. Hirano, and T. C. Chiles. 1996. Transcriptional regulation of the junB promoter in mature B lymphocytes. Activation through a cyclic adenosine 3′,5′-monophosphate-like binding site. J. Immunol. 157:146-155. - PubMed
-
- Arthur, J. S. C., and P. Cohen. 2000. MSK1 is required for CREB phosphorylation in response to mitogens in mouse embryonic stem cells. FEBS Lett. 482:44-48. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials