

The intraflagellar transport protein, IFT88, is essential for vertebrate photoreceptor assembly and maintenance
- PMID: 11916979
- PMCID: PMC2173265
- DOI: 10.1083/jcb.200107108
The intraflagellar transport protein, IFT88, is essential for vertebrate photoreceptor assembly and maintenance
Abstract
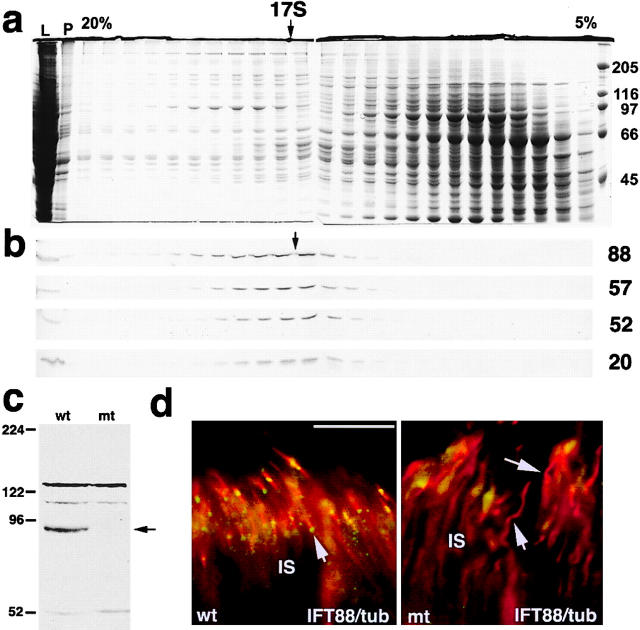
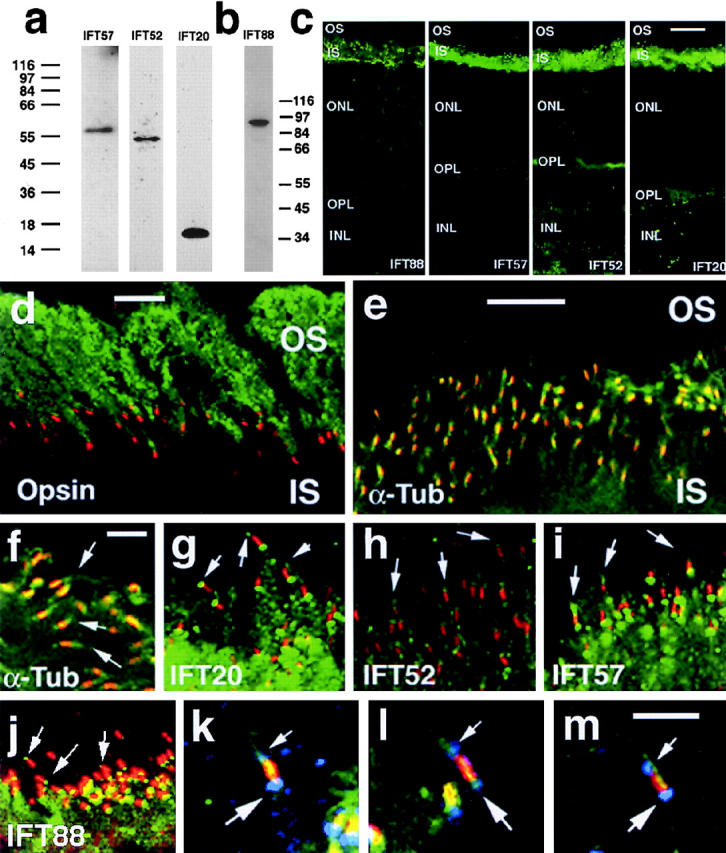
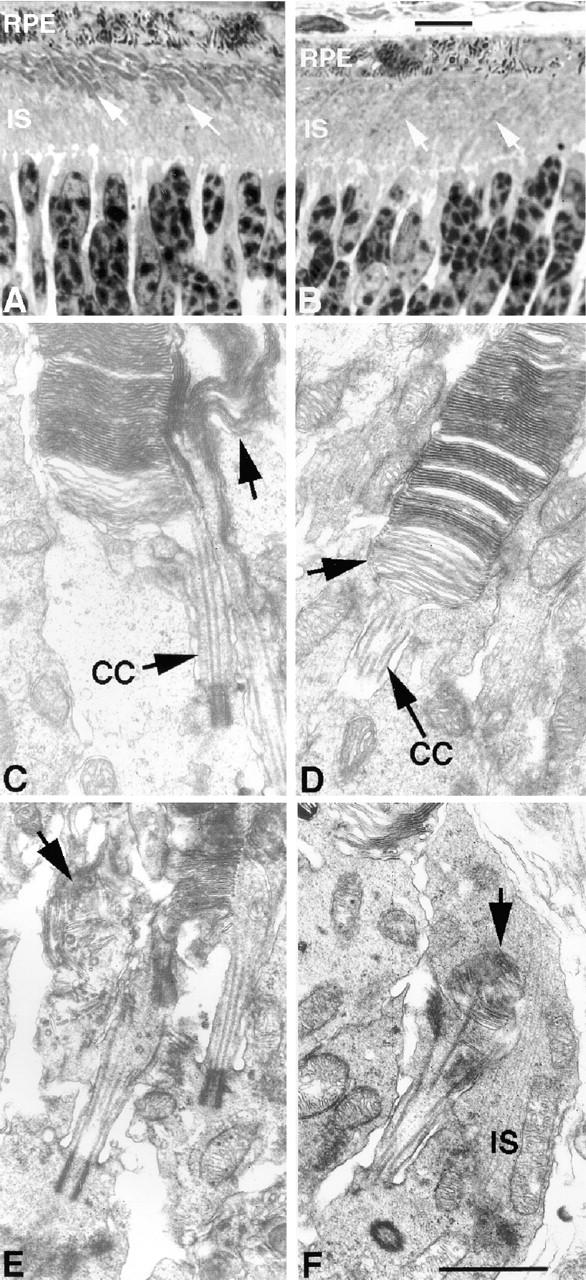
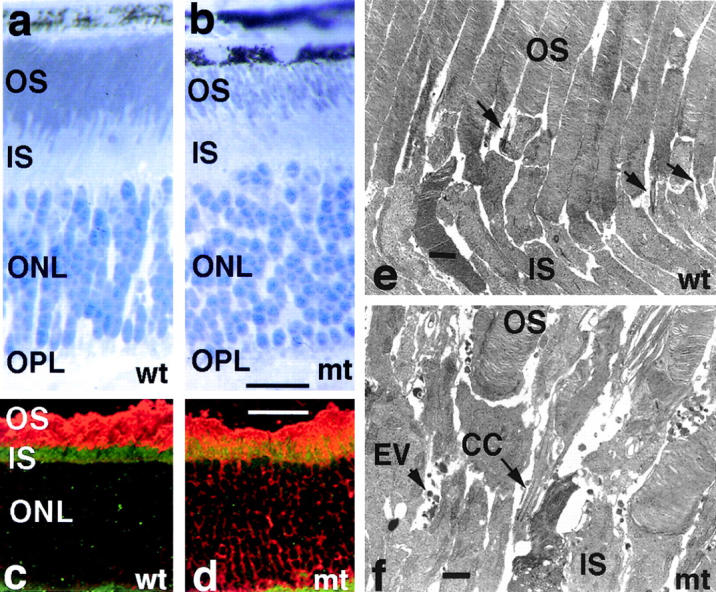
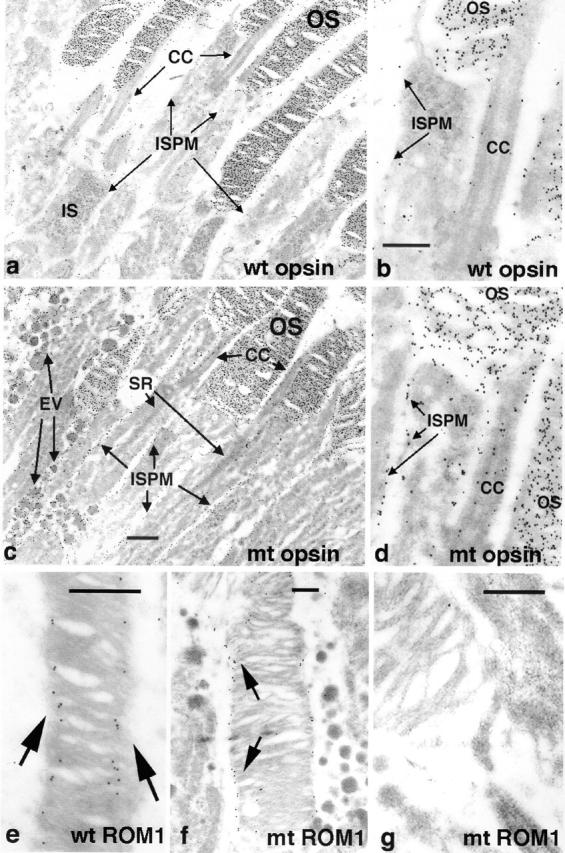
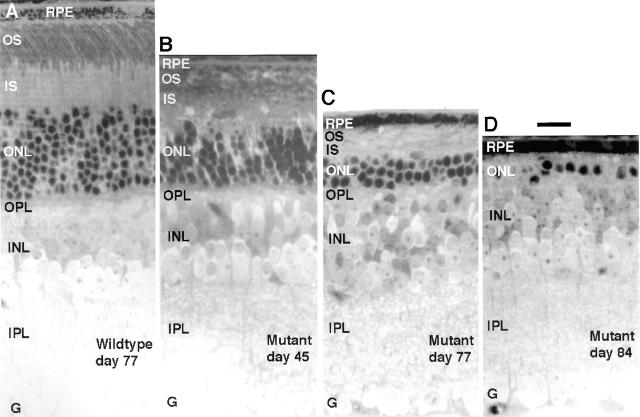
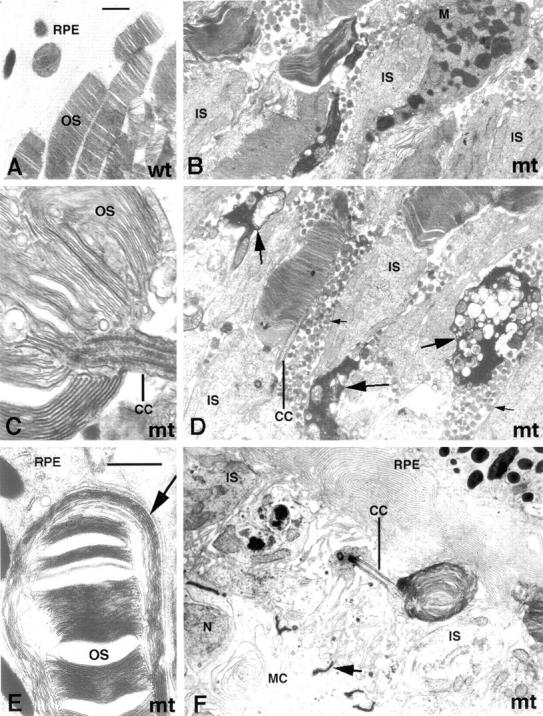
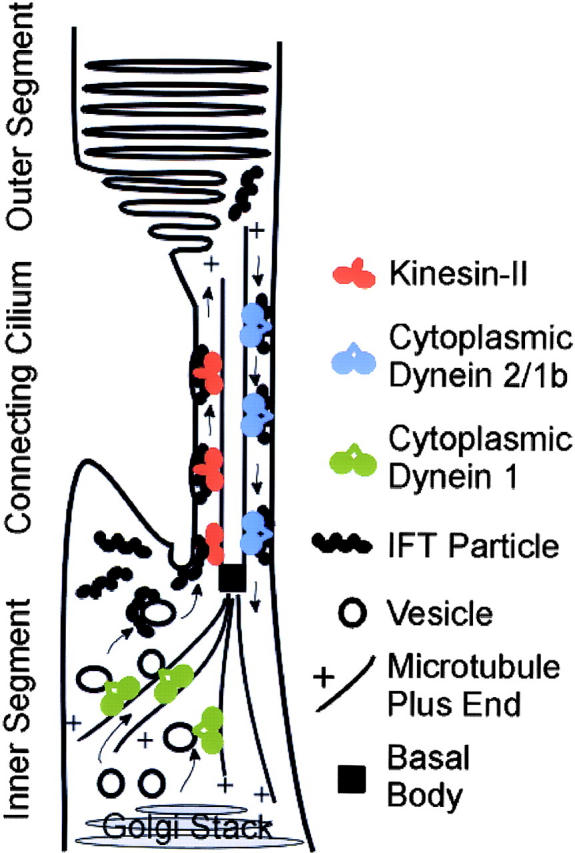
Approximately 10% of the photoreceptor outer segment (OS) is turned over each day, requiring large amounts of lipid and protein to be moved from the inner segment to the OS. Defects in intraphotoreceptor transport can lead to retinal degeneration and blindness. The transport mechanisms are unknown, but because the OS is a modified cilium, intraflagellar transport (IFT) is a candidate mechanism. IFT involves movement of large protein complexes along ciliary microtubules and is required for assembly and maintenance of cilia. We show that IFT particle proteins are localized to photoreceptor connecting cilia. We further find that mice with a mutation in the IFT particle protein gene, Tg737/IFT88, have abnormal OS development and retinal degeneration. Thus, IFT is important for assembly and maintenance of the vertebrate OS.
Figures
References
-
- Anderson, R.E., M.B. Maude, P.A. Kelleher, T.M. Maida, and S.F. Basinger. 1980. Metabolism of phosphatidylcholine in the frog retina. Biochim. Biophys. Acta. 620:212–226. - PubMed
-
- Beech, P.L., K. Pagh-Roehl, Y. Noda, N. Hirokawa, B. Burnside, and J.L. Rosenbaum. 1996. Localization of kinesin superfamily proteins to the connecting cilium of fish photoreceptors. J. Cell Sci. 109:889–897. - PubMed
-
- Besharse, J.C., and C.J. Horst. 1990. The photoreceptor connecting cilium. A model for the transition zone. Ciliary and Flagellar Membranes. R.A. Bloodgood, editor. Plenum Publishing Corp, New York. 389–417.
-
- Besharse, J.C., and M.G. Wetzel. 1995. Immunocytochemical localization of opsin in rod photoreceptors during periods of rapid disc assembly. J. Neurocytol. 24:371–388. - PubMed
-
- Blanks, J.C., and C. Spee. 1992. Retinal degeneration in the pcd/pcd mutant mouse: accumulation of spherules in the interphotoreceptor space. Exp. Eye Res. 54:637–644. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
- GM-30626/GM/NIGMS NIH HHS/United States
- GM-14642/GM/NIGMS NIH HHS/United States
- GM-60992/GM/NIGMS NIH HHS/United States
- R01 GM030626/GM/NIGMS NIH HHS/United States
- EY-03222/EY/NEI NIH HHS/United States
- R37 GM030626/GM/NIGMS NIH HHS/United States
- R01 EY003222/EY/NEI NIH HHS/United States
- P30 DK032520/DK/NIDDK NIH HHS/United States
- R01 GM014642/GM/NIGMS NIH HHS/United States
- R37 GM014642/GM/NIGMS NIH HHS/United States
- EY01931/EY/NEI NIH HHS/United States
- DK 32520/DK/NIDDK NIH HHS/United States
- R01 GM060992/GM/NIGMS NIH HHS/United States
- T32 EY014537/EY/NEI NIH HHS/United States
- P30 EY001931/EY/NEI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
