

The procaspase-8 isoform, procaspase-8L, recruited to the BAP31 complex at the endoplasmic reticulum
- PMID: 11917123
- PMCID: PMC123648
- DOI: 10.1073/pnas.072088099
The procaspase-8 isoform, procaspase-8L, recruited to the BAP31 complex at the endoplasmic reticulum
Abstract
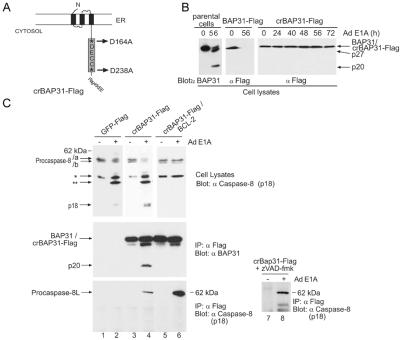
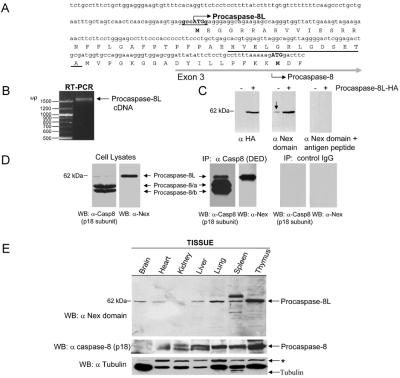
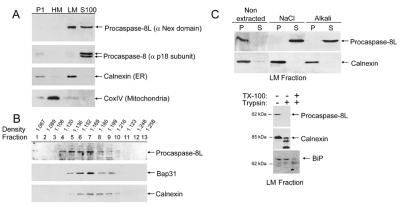
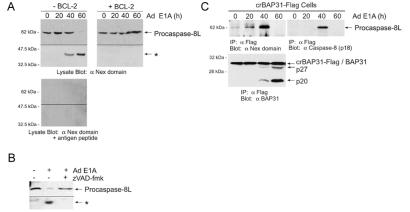
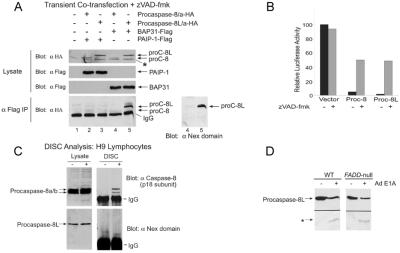
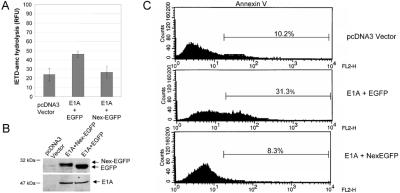
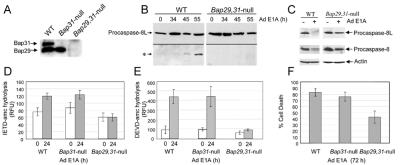
BAP31 is an integral protein of the endoplasmic reticulum membrane and a substrate of caspase-8. Here, we describe the procaspase-8 isoform, procaspase-8L, which is ubiquitously expressed and selectively recruited to the BAP31 complex in response to apoptotic signaling by E1A. Procaspase-8L is characterized by the N-terminal extension (Nex) domain, which extends procaspase-8/a at the N terminus and is required for selective association of procaspase-8L with the BAP31 complex. Gene deletion identified BAP31 and related BAP29 as required for processing of procaspase-8L in response to E1A, by a FADD-independent mechanism that was blocked by BCL-2. Further, Bap29,31 deletion, as well as a Nex-domain dominant-negative mutant, curtailed the activation of downstream caspases (IETDase and DEVDase) and cell death in response to E1A. Preferential recruitment of procaspase-8L by the BAP31 complex at the endoplasmic reticulum suggests an additional pathway for regulating initiator caspase-8 during apoptosis.
Figures
References
-
- Budihardjo I, Oliver H, Lutter M, Luo X, Wang X. Annu Rev Cell Dev Biol. 1999;15:269–290. - PubMed
-
- Earnshaw W C, Martins L M, Kaufmann S H. Annu Rev Biochem. 1999;68:383–424. - PubMed
-
- Korsmeyer S J, Wei M C, Saito M, Weiler S, Oh K J, Schlesinger P H. Cell Death Differ. 2000;12:1166–1173. - PubMed
-
- Mosser J, Sarde C O, Vicaire S, Yates J R, Mandel J L. Genomics. 1994;22:469–471. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
